獣医生理学教室では哺乳類における個体および種の維持機構の解明という観点から、「成長と生殖の分子機構」をメインテーマに、神経生物学的あるいは細胞生物学的研究手法を取り入れた幅広い研究を展開しています。研究には私達自身で作出した遺伝子改変動物(遺伝子を適当なプロモーターと組み合わせて導入したトランスジェニックラットや、特定の遺伝子を破壊したノックアウトマウス)を多用するとともに、附属牧場で系統造成されているシバヤギ(研究用のクローズドコロニーの確立している在来種)等を用いた実証研究にも取り組んでいます。動物体内で営まれる生命現象とその調節機構に関する基礎的研究を基盤として、新しい動物産業への展開や病態に対するより深い理解など、応用面への貢献を目指すことができればと考えています。具体的には、骨格筋の発達・再生に関する研究と中枢神経系の働きに関する研究の2つを柱としています。
骨格筋の発達・再生に関する研究
骨格筋は多核の筋線維から構成されており、生体全体の40%を占める体内で最大の組織です。筋線維は損傷しても速やかに再生することができます、これは骨格筋に存在する筋衛星細胞という組織特異的幹細胞(前駆細胞)が常に一定の割合で存在するためです。
筋損傷時におこる筋衛星細胞による筋線維の再生は、間葉系前駆細胞や免疫系の細胞、血管内皮細胞など、他の細胞群と相互作用することで達成されます。
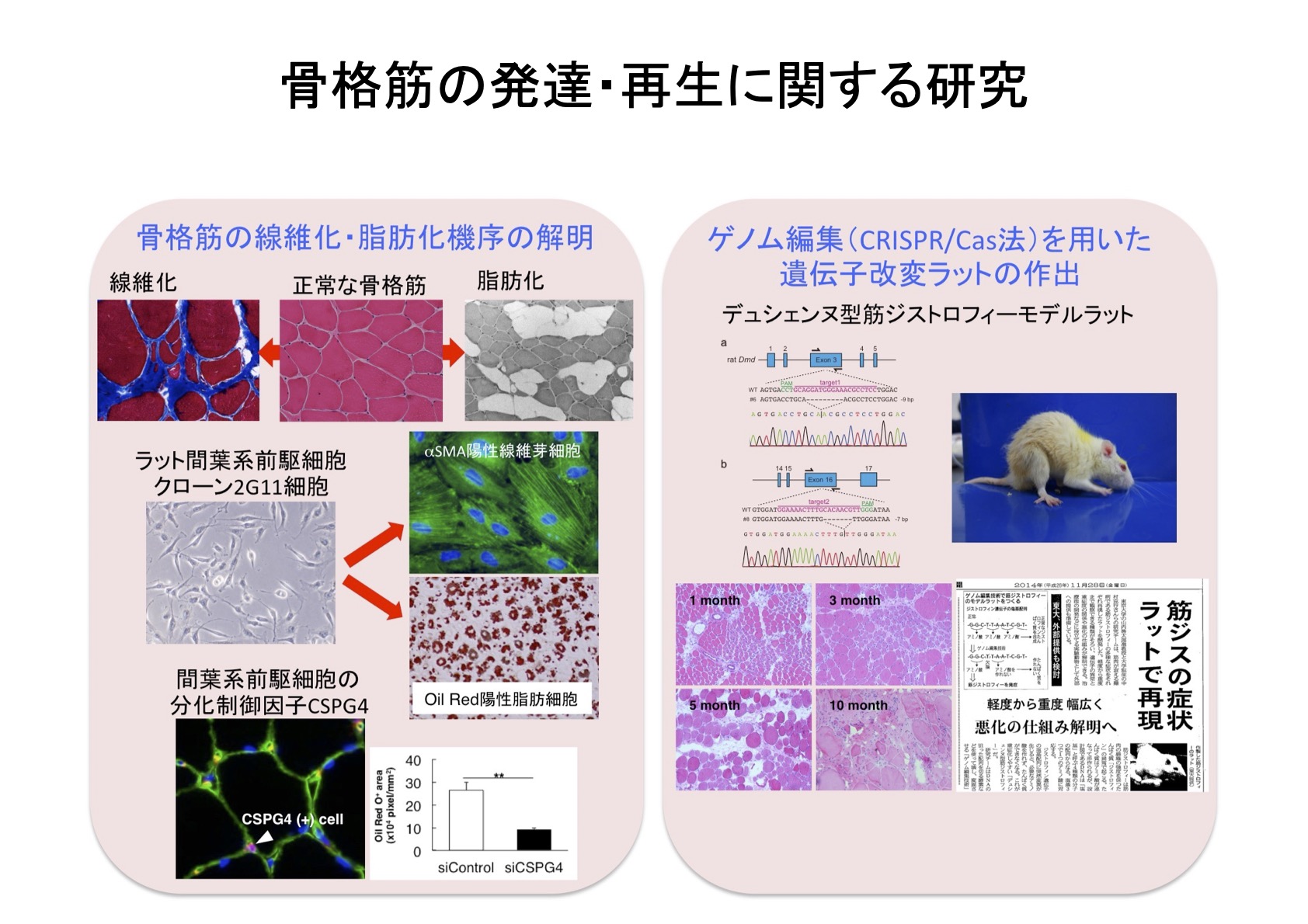
一方、ヒトの重篤な遺伝性筋疾患であるデュシェンヌ型筋ジストロフィー(DMD)では、筋線維の損傷と再生が繰り返され、最終的には筋再生が追いつかないために患者は最終的に呼吸不全により死に至ります。
私たちの教室では、ヒトDMDの病態進行をよく反映したDMDラットの作出に世界で初めて成功しました(Nakamura et al., 2014)。DMDラットを用いたこれまでの研究から、DMDでは早期細胞老化という減少が生じておりこれがDMDの病態悪化に大きく関わっていることを見出しました(Sugihara et al., 2020)。
DMDやサルコペニアのような筋疾患では、骨格筋内に線維化(線維芽細胞による間質への過剰なコラーゲンの沈着)や脂肪化(間質への脂肪細胞の出現)がみられます。この線維化や脂肪化はいずれも筋機能を著しく阻害するだけでなく、筋再生能の低下を招きます。線維化や脂肪化は間葉系前駆細胞が線維芽細胞や脂肪細胞へと分化することでおこります。私たちはラット間葉系前駆細胞のクローン化に成功し、その細胞(2G11細胞)を利用してこれまでに間葉系前駆細胞の分化制御機構について明らかにしてきました(Takeuchi et al., 2020)。
これまで骨格筋の発生・成長・再生に関する研究には広くマウスが用いられてきましたが、私たちの研究によりマウスで得られてきた知見は必ずしもヒトや他の動物へと敷衍できないことが判明しています。そのためこれまで教科書に記載されているような事項についてもあらためて比較生物学的視点から再検証する必要があると考えています。
以上を背景として、現在は、以下のようなprojectを中心に進めています。
1)遺伝子改変ラットを利用したDMDなどの筋疾患の病態成立機序の解明
2)間葉系前駆細胞の分化制御機構の解明
中枢神経系の働きに関する研究
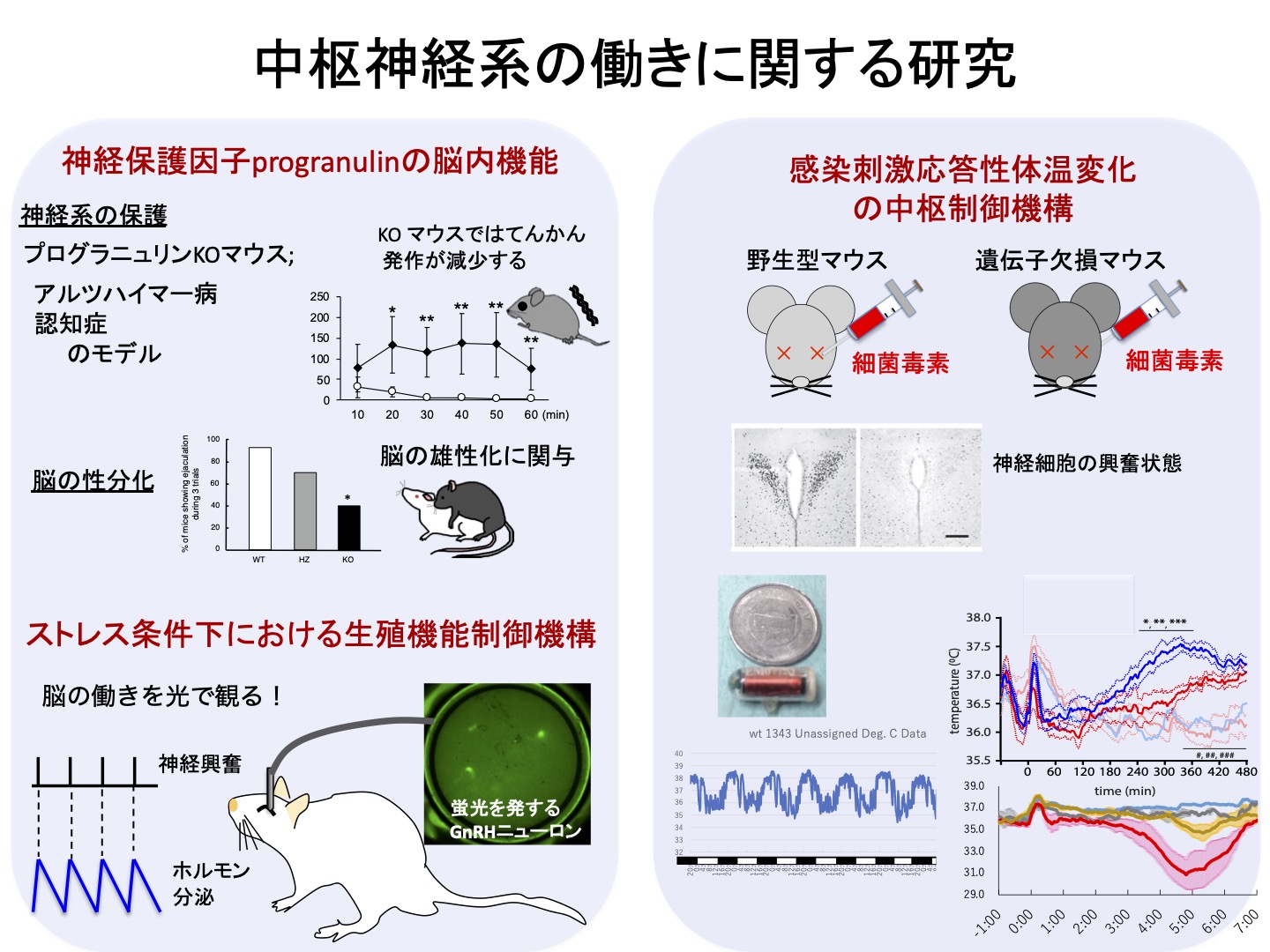
神経保護因子progranulinの脳内機能
哺乳類の脳には組織学的・機能的な雌雄差が存在します。発生・成長過程において、脳が未分化な状態から雄型もしくは雌型に発育していくことを、脳の性分化と呼びます。我々の研究室では、脳の性分化における重要な因子の一つとしてプログラニュリン(Progranulin, PGRN)を同定し、その脳内作用機序について研究を続けています(Suzuki et al., 1998; Kayasuga et al., 2007他)。一方で近年、他のグループの研究によってPGRNの遺伝子異常が前頭側頭葉変異症やアルツハイマー病などの神経変性性疾患の原因となることが明らかとなり、PGRNの神経保護的な役割が世界的に注目されています。我々もこれまでの研究に加えて、中枢神経系の病態発現やその抑制に関連したPGRNの働きについて研究を行っており、最近の成果としてPGRNがてんかん発作などの神経細胞の異常興奮を抑制する働きをもつことを明らかにしました(Kuroda et al., 2020)。
ストレス条件下における生殖機能制御機構
哺乳類の生殖機能は神経内分泌的に、間脳視床下部から分泌される性腺刺激ホルモン放出ホルモン(GnRH)、GnRHの刺激を受けて下垂体から分泌される黄体形成ホルモン(LH)と卵胞形成ホルモン(FSH)によって制御されています。GnRHは雌雄に共通して存在するパルス状と雌特有のサージ状という二つの分泌様式をもち、雌の動物では卵巣にLHとFSHがパルス状に作用することで卵胞を発育させ、その後LHがサージ状に働くことで排卵が誘起されます。私たちの研究室では長年、自由行動下の動物の視床下部の電気生理学的な観察(Yoo et al., 1997他)や頸静脈に留置したカニューレからの連続採血(Matsuwaki et al, 2003, 2006他)によってGnRHやLHのパルス状分泌の経時的な観察を続けてきました。このGnRH/LHパルスを生殖機能の指標としながら、様々なストレス条件下における生殖機能制御機構について、中枢神経系の働きを中心に解析を進めています。その過程で、ストレス条件下で時にGnRH/LHパルスすなわち生殖機能を抑制するのは、脳内で増加したプロスタグランジン類であることを明らかにしました(Matsuwaki et al., 2017)。さらに現在は、GnRHニューロン特異的に蛍光を発するラットの脳内に蛍光顕微鏡に接続した光ファイバーを刺入することで、GnRHニューロンの即時的な蛍光観察を試みています。
感染刺激応答性体温変化の中枢制御機構
哺乳類の体温は通常一定の範囲で恒常性をもって保たれていますが、細菌やウイルスなどへの感染により、その範囲を越えた上昇もしくは低下が起きます。このような体温調整の中心は間脳の視床下部であると考えられています。感染性の体温上昇については古くから研究が行われており、感染時に脳内で増加したプロスタグランジンE2が熱源物質として働くことがよく知られています。我々のグループではこれまで、末梢組織の感染情報によって血中で増加した炎症性サイトカインが脳内の血管内皮細胞に作用し、同細胞で産生されたプロスタグランジンE2が視床下部で働くことで体温が上昇することを見出しました(Matsuwaki et al., 2014 & 2017; Eskilsson et al, 2017他)。一方で、感染性体温低下の詳細な機構については未だ不明な点が多く残されています。我々の研究グループは、熱源物資であるプロスタグランジンE2の合成酵素の遺伝子を欠損するマウスの一部では、感染刺激時に体温が上昇しないだけではなく強度の体温低下を呈することを見出しました。さらに、その他いくつかの低体温発症モデルを用いて、感染性低体温の発症機構についての研究を進めています。