研究紹介
マウスの器官形成におけるMAB21の機能解析
◯主な研究成果(クリックすると各項目のトップに移動します)
- 1. シングルセルRNAseq解析によるマウス眼胞の網膜前駆細胞のHeterogeneityとその分子基盤
- 2. MAB21L1はレンズ前駆細胞での遺伝子発現とDNA代謝経路を制御する
- 3. ヌクレオチド代謝関連因子MAB21L1は末梢の急速な表層外胚葉の形態形成においてエネルギー代謝的にサポートする
-マウス外生殖器形成を例に-
1. シングルセルRNAseq解析によるマウス眼胞の網膜前駆細胞のHeterogeneityとその分子基盤
株) RNA - 共同研究プロジェクト費「シングルセルRNA解析によるMAB21機能研究」哺乳類の眼の網膜原基である眼胞(optic vesicle)は、前脳予定領域の神経外胚葉から連続する左右一対の憩室として発生する。眼胞の網膜前駆細胞への運命決定は、4種類のマスター制御因子EFTFs (eye field transcription factors; PAX6, SIX3, RAX, LHX2) が互いに協調して、その下流に位置するMAB21L2 (MABドメイン因子の1つ)を含めた網膜関連因子の発現を誘導し、眼杯と呼ばれる網膜構造が形成される。今まで眼杯からの様々な網膜の構成細胞への分化の解析は詳細になされてきたが、残念ながら初期の眼胞の網膜前駆細胞の遺伝子の発現変動は不明であり、この時期の網膜前駆細胞のheterogeneityの存在も明らかにされていない。私たちは、網膜前駆細胞でのEFTFs直下の遺伝子カスケードとHeterogeneityの分子基盤を明確にするため、胎齢8.5-9.5日を6時間ごとの5ステージ(12-, 16-, 20-, 24-, 26-somite stages)の眼胞(合計7,989 cells)からシングルセルRNA解析を行い、網膜前駆細胞(PAX6/SIX3/RAX-positive細胞) に焦点を当て、網羅的な単一細胞レベルの遺伝子発現プロファイルを作成した(下図)。
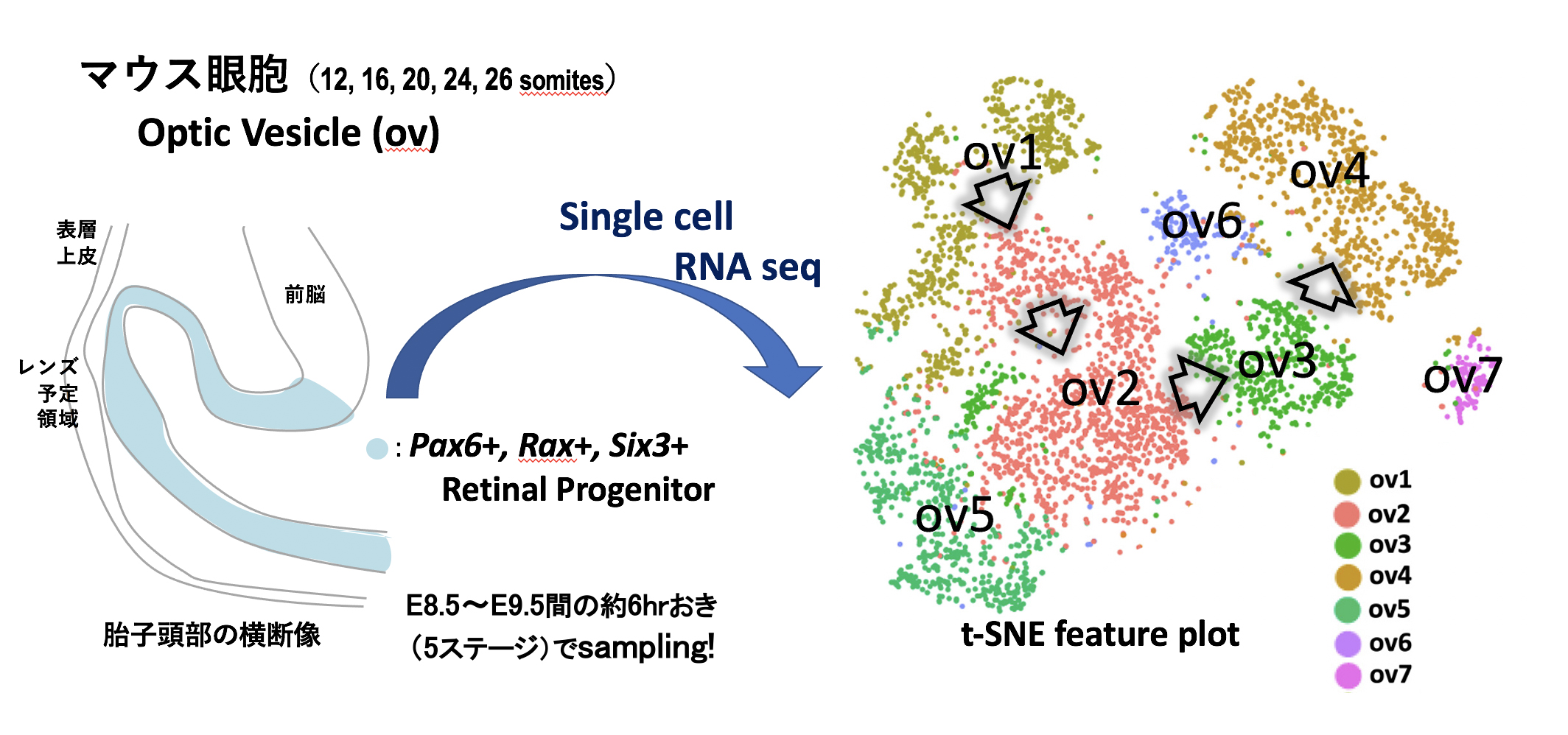
tSNE (T-distributed stochastic neighbor embedding) plot により、5ステージ由来の眼胞内の網膜前駆細胞は7種類のクラスター(ov1~ov7)に分類することができた。このクラスター内で ov1, ov2, ov3, ov4は発生ステージ特異的なクラスターであり、ov1は約80%が12-somite由来、ov2は約94%が16-18-somite由来、ov3およびov4は95%以上の細胞が各々24-および26-somite由来の網膜前駆細胞であった(上図右参照:矢印の方向へと分化が進行)。各クラスターのDEGs (differentially expressed genes) 解析の結果、1)ov1はWnt8b+の未分化な神経外胚葉の特性を維持しており、2)ov2以降へと発生ステージが進むに連れてCol9a1, Vim, Ckb などの細胞骨格、細胞外マトリックス、エネルギー代謝系の遺伝子の発現が上昇し、3)ov4から EFTFs直下のMab21l2, Tbx5, Vax2, Pax2 etc. の網膜の各種細胞への分化に関わる鍵遺伝子の発現が上昇することを単一細胞レベルで証明することができた。なお、上記以外のov5-ov7のクラスターに関しては、ov6, ov7に含まれる網膜前駆細胞は、全発生ステージに存在する Malat1(non-coding RNAの一つ)-negativeのサブタイプであり、一方、ov5はPax6低発現の前脳との境界部に位置するサブタイプであると想定された。
本成果は、哺乳類の眼胞内の網膜前駆細胞のheterogeneityの存在を単一細胞レベルで分類し、さらに発生段階に依存した遺伝子発現変動を明確にした最初の報告となった。眼胞の網膜前駆細胞は、26-somite(ov4に相当)から数時間後から眼杯へのダイナミックな形態形成を開始する。得られた発生段階に伴った網膜前駆細胞の代謝・構造関連の遺伝子変動は、眼胞期の眼杯への陥入運動の“準備状態“を深く反映しているものと想定された。(Yamada et al., BBRC, 2021)。
2. MAB21L1はレンズ前駆細胞での遺伝子発現とDNA代謝経路を制御する
株) RNA - 共同研究プロジェクト費「シングルセルRNA解析によるMAB21機能研究」哺乳類の眼のレンズ原基である水晶体板(lens placode)は、前脳予定領域の左右一対の眼胞と接する表層外胚葉から誘導される。この水晶体板の形成には3種類のマスター制御因子 SIX3, PAX6, SOX2が互いに協調して、その下流に位置するMAB21L1 (MABドメイン因子の1つ)などのレンズ形成関連因子の発現を誘導する。水晶体板の中央部分からレンズが陥入し、表層に残された外胚葉表皮からは虹彩、角膜が誘導される。MAB21l1は、ヒトCOFG(congenital cerebellar, ocular, craniofacial, and genital)症候群の原因遺伝子であり、その遺伝子変異は、レンズ、虹彩、角膜を含めた眼の前眼部、陰嚢(マウスでは陰嚢領域の包皮腺)が低形成となる。MAB21l1の発現パターンとその欠損(KO)胎子の表現型から、形態形成時のエネルギー供給(血管分布)の乏しい末梢組織での細胞死を特徴とすることから、MAB21L1は低エネルギー状態下での形態形成に機能していると想定される。しかし、残念ながらMAB21L1の生理学的な機能は不明である。
私たちは、Mab21l1 KOマウス由来のレンズ前駆細胞を用いてsingle cellレベルでの電顕病理、発現解析を行った。その結果、水晶体板では、MAB21L1は核内に存在し(核内で機能し)、Mab21l1 KO水晶体板は、表層の糖衣(Glycocalyx)の成熟(肥厚)が進行せず、GRP78/BiPの蓄積を伴う小胞体ストレスが細胞死の直前に誘導されることが判明した。次に、KO水晶体板で細胞死の起こる直前(26-somite stages)でのシングルセルRNA(scRNA)解析を行い、網羅的な単一細胞レベルの遺伝子発現プロファイルを作成した(下図A, 水晶体板の模式図; 図B, scRNA-seqのtSNE plot)。そこから、レンズ前駆細胞(Epcam/Krt8/Cldn3-positive上皮細胞)のみを抽出し、野生型とKO細胞間の発現の異なる遺伝子変化を解析した。その結果、野生型と比べてKO細胞において、レンズ特異的な21遺伝子を含む131遺伝子が低下し、表層上皮特異的な22遺伝子を含む148遺伝子が上昇していることを見出した。これら変動した遺伝子(DEGs)のGO解析の結果、アポトシース関連遺伝子の変動と共に、DNA代謝、複製に関与する遺伝子が減少、有糸分裂に関与する遺伝子群が上昇していることが判明した(下図C)。
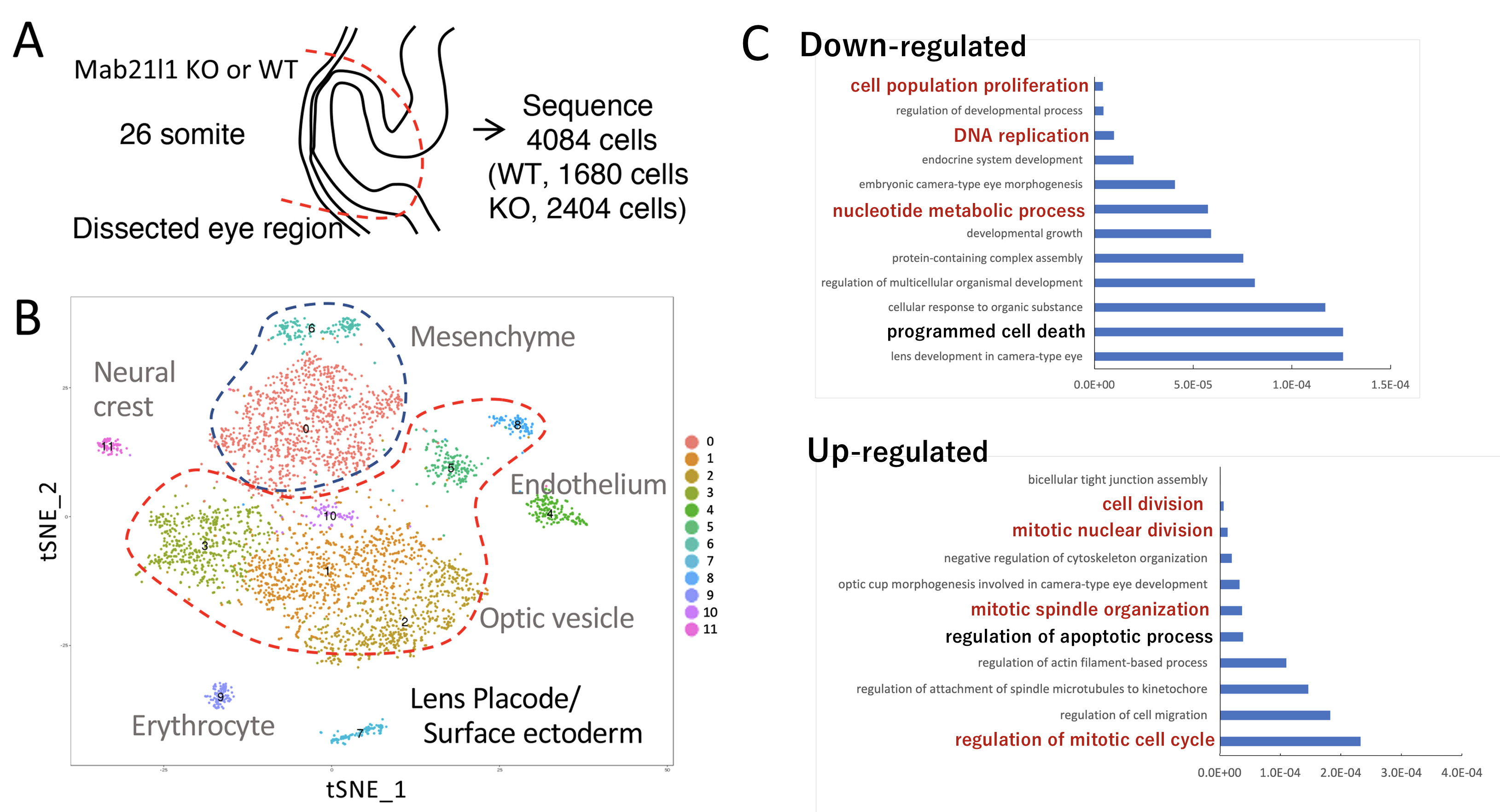
本研究により、MAB21L1は、LP細胞でのDNA代謝・複製に関与する遺伝子群の一連の発現を正常レベルに維持するのに必須であることが明らかとなった。別の有名なMABドメイン因子であるcGAS(環状GMP-AMPシンターゼ)は、MAB21L1と類似したタンパクの高次構造をとるが、cGASはATP/GTPと結合し、これらを消費することが知られている。一方、MAB21L1の方は、CTPと結合し、この結合部位はMAB21L1の機能に重要であることが判明している。おそらく、MAB21L1は、cGASのATP/GTPの代謝と相反する作用方向で、CTP結合部を介してDNA代謝を維持し、低エネルギー状況下での糖衣(Glycocalyx)合成 etc.の成熟過程や続く水晶体板の形態形成をエネルギー的に支持しているものと推測される(Yamada et al., Disease Model Mech., 2021)。
3. ヌクレオチド代謝関連因子MAB21L1は末梢の急速な表層外胚葉の形態形成においてエネルギー代謝的にサポートする -マウス外生殖器形成を例に-
株) RNA - 共同研究プロジェクト費、笹川科学研究助成(2022-4106)Mab21l1(Male abnormal 21-like 1)は、核酸代謝に関与するヌクレオチジルトランスフェラーゼ(NTase)フォールドタンパク質ファミリーのメンバーである。 ヒトMAB21L1変異は、小脳-眼-頭蓋顔面-外性器(COFG)症を引き起こし、男児、女児共に外陰部の低形成を呈する。Mab21l1欠損マウスも、小眼球症や頭蓋骨形成不全とともに、 陰嚢ヒダに由来する齧歯類特異的な包皮腺の低形成が誘導される。 これらの異常は、血液供給が乏い末梢組織の形態形成の部位であり、 かつその表現型も支配血管から遠い部位でsevereな表現型を呈することから、おそらくMAB21L1は、血液供給の乏しい末梢組織の形態形成をエネルギー的にサポートしているものと推測される。 しかし、MAB21Llが、マウス、ヒトの最も末梢の表層外胚葉外性器のどこで、どのように機能しているのかは不明であった。 我々は、マウス外性器の形態形成におけるMAB21L1の役割を明確することを目的として、Mab21l1-/-新生子マウスの外性器の表現型の解析を行なった。 その結果、1)Mab21l1-/-新生子マウスは、ヒトCOFG症と同様に、陰嚢、包皮腺の外胚葉上皮の低形成を呈し、包皮腺の腺上皮の増殖の低下、 グリコーゲン蓄積の低下(低栄養状態)を示した。2) MAB21L1の発現部位は、この上皮の裏打ちする間葉系細胞の一部に強く発現していること(特殊なMAB21L1+間葉系細胞の存在)を見出した。 そこで、これらのMab21l1-/-包皮腺を裏打ちする間充織の詳細な表現型解析の結果、3)MAB21L1+間葉系細胞の細胞密度、増殖率の有意な低下、 4)細胞外基質(ECM;コラーゲンI、フィブロネクチン、ヘパラン硫酸)の減少が判明した。さらに、5)胎齢18.5日の野生型とMab21l1-/-の包皮腺原基の器官培養での解析により、 Mab21l1-/-の腺構造は、野生型と比べ低形成を示し、マトリゲル(ECM mixture)の添加により、腺構造の形成、上皮の細胞増殖活性は野生型と同レベルまで回復することが判明した。 以上の結果から、外陰部の形態形成では、MAB21L1+ 間葉系細胞は、低エネルギー状況下での自己増殖、ECM産生を可能にし、末梢領域での上皮の形態形成をサポートしていることが推測された。

以上の結果から、水晶体板etcと同様に、外陰部でも、「MAB21L1は、そのヌクレオチド代謝調節により、末梢の表層外胚葉の素早い形態形成をエネルギー代謝的にサポートする」 という仮説を支持するものであった。MAB21L1/2は、同じMABドメイン因子であるcGAS(環状GMP-AMPシンターゼ)に対してドミナントネガティブに作用できるタンパク構造を示すことから、 このヌクレオチド代謝調節の分子機序は、cGASを抑制することにより働くのか、あるいは別の制御があるのかについては今後の解析が必要となる (Promsut et al. The Anatomical Record, 2023)。
