研究内容
研究計画・方法
腸上皮基底膜直下にシート状に分布する腸筋線維芽細胞、類洞細胞を覆う肝星細胞(伊東細胞)、肝小葉間隙を埋める肝筋線維芽細胞、腸管外側を包む中皮細胞、腸管運動ペースメーカー機能をもつc-kit 陽性カハール介在細胞、そしてPDGFRαとSK3 を発現するがc-kit 陰性の線維芽細胞様細胞(2009 年に研究分担者飯野により新規に同定された間質細胞)を研究対象とする。特に着目する分子は、SRF/myocardin(転写制御因子)、CPI-17 とMYPT-1(筋収縮制御因子)、tenascin C(細胞遊走の軌道となる分子)、aquaporin-1(細胞変形をになう分子)などである。
平成25 年度の研究計画
【項目1】筋線維芽細胞の筋分化機構(転写制御系の解明)
血清応答因子Serum Response Factor (SRF) は、平滑筋細胞に特異的な遺伝子(αSMA、SM22、平滑筋ミオシン重鎖など)のプロモーター中に存在するシス配列(CArG ボックス)に結合して転写を促進する。Myocardin はこのSRF の活性化補助因子として見出されたものである。本項目では間葉系細胞における収縮能獲得のメカニズムを解明する。
1) 筋蛋白系発現転写制御とSRF/myocardin 系の関係の関係
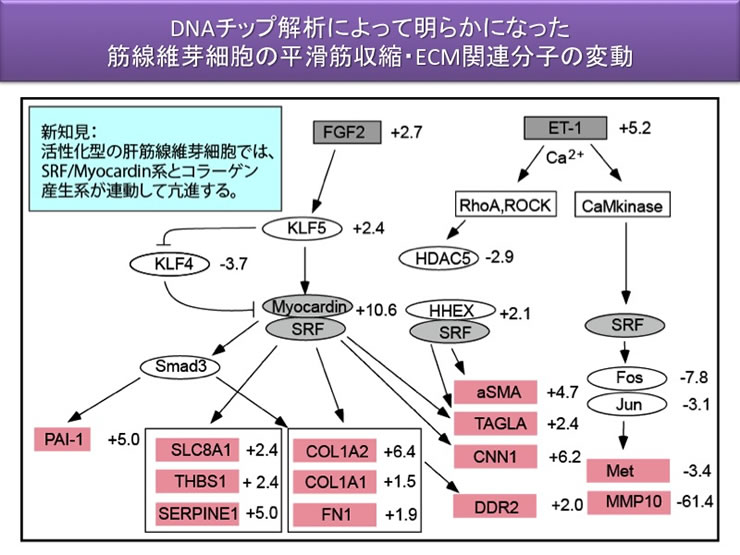
すでにDNA チップ解析結果から、肝筋線維芽細胞の活性化に伴って、αSMA の発現増加以外にもカルポニンやカルデスモン、CPI-17 などの筋蛋白関連遺伝子群の発現がmRNA レベルで上昇する成績を得ており(図2)、Real-time RT-PCR による確認もしている。筋蛋白系の他の分子群として、カルポニン、カルデスモンなどのアクチン結合蛋白質や内因性脱リン酸化酵素阻害蛋白質CPI-17 やMYPT-1 の発現変動について解析する。
2) 収縮機能と線維素産生機能の連関機構
予備研究において、肝星細胞にmyocardin のsiRNA 処置した活性型肝筋線維芽細胞ではαSMAの発現量が低下していただけでなく、予想に反してコラーゲン遺伝子の発現も低下していることを突き止めている。本項目では、この点についてさらに蛋白質レベルでの検証を行うとともに、SRF のsiRNA、myocardin の強制発現などの細胞工学的実験を追加し、筋蛋白質発現系とコラーゲン産生系の転写活性が連関するメカニズムを解明する。
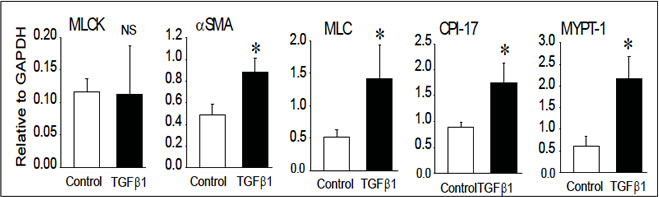
★図2 重要な予備データ(1):
ラット腸上皮細胞をTGFβ1 で7 日間処理してEMT を惹起した。リン酸化酵素であるMLCK は変化しないが、αSMA、ミオシン軽鎖、CPI-17、MYPT-1 などの収縮蛋白発現が上昇していることが確認された。SRF/Myocardin 系制御が強く疑われる。
【項目2】収縮機能情報伝達系の解明
収縮機能が細胞内カルシウム濃度上昇をともなうミオシンリン酸化機構で説明できるか、CPI-17 やMYPT-1 などが関与する脱リン酸化制御機構は存在するかなどを明らかにする。肝星細胞における脱リン酸化制御機構に関する基礎的知見を得ているが、関連蛋白質の発現制御(siRNAによる抑制)を行うことによりその詳細を解明する。
1) 細胞遊走活性と収縮機能
円形のコラーゲンゲル上に細胞を播種し、その径の変化を観察することにより活性型と非活性型の筋線維芽細胞の収縮を計測する。またボイデンチャンバーを使ったmigration assay 法あるいはwound healing assay 法を用いて、細胞遊走について解析を行う。転写制御系の解明にで得られた収縮蛋白系の分子発現についての情報を利用し、細胞遊走に重要な細胞内情報伝達機能について解析する。
2) ミオシンリン酸化機構の解析
PKC、Rho キナーゼなどを介するCPI-17 あるいはMYPT-1 のリン酸化にともなうミオシン軽鎖の脱リン酸化の制御機構について詳細に検討する。
【項目3】筋線維芽細胞のソースとしての上皮・間葉転換(EMT)
上皮細胞に比べて間葉系細胞は極性に乏しく高い遊走能をもつ。一般に上皮はE-cadherin を発現しているのに対して,間葉系細胞はN-カドヘリン,R- cadherin やcadherin -11 を発現している。本項目では、TGFβ で誘導される上皮・間葉転換において、E-cadherin の消失と筋分化がどの様に連関しているかを解析する。
【項目4】筋線維芽細胞が産生する障害修復因子としての細胞外マトリックスの役割
Tenascin-C は1986 年に発生期に特異部位に出現する糖蛋白として発見され、その後の研究で各種の炎症や腫瘍形成に関わることが明らかにされた。産生細胞は平滑筋や線維芽細胞をはじめとする間葉系細胞とされているが、細胞内で合成されると直ぐに細胞外へ放出され周囲に沈着することから、詳しい研究が行われてこなかった。Tenascin-C は単なるマトリックス蛋白質ではなく、TRL4 受容体の補助活性化因子となることが明らかになるなど(Midwood ら, Nature Med 2009)、機能分子としての役割が注目されている。
予備知見として、腸管上皮細胞が障害後に上皮・間葉転換する際に、tenascin-C を産生し、オートクラインに働いて細胞遊走を活性化する現象を見いだしている(図3)。
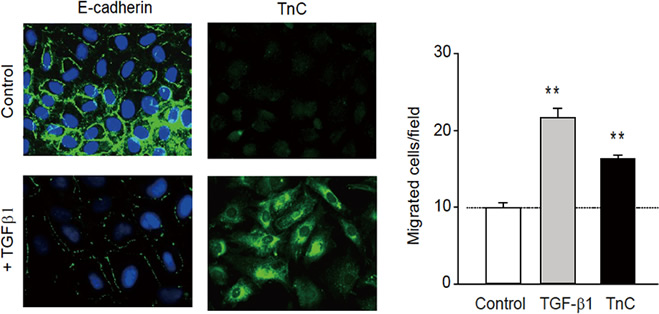
★図3 重要な予備データ(2):腸上皮細胞をTGFβ で刺激してEMT を起こさせるとE-Cadherin が減少しtenascin-C が上昇する(右)。Transwell-well assay で上皮細胞の遊走活性を測るとTenascin-C をコーティングすることで増加する。
【項目5】細胞遊走時の細胞変形の分子機構(水チャネルの役割)
細胞が遊走を開始する時には、細胞先導端でアクチンが重合することが知られている(Lauffenburger ら, Cell 1996)。しかし、重合するための空間がどのように形成されるのかについては不明である。Saadoun ら(Nature 2005)は、細胞が遊走する直前には、遊走細胞の先導端に水チャネルであるアクアポリン分子が局在し、そのために局所的な水流入と膨化が生じて先導端にアクチンが重合できる空間が形成されることを観察し、アクチン重合のための空間確保の分子機序の理解が進んだ。本項目では、細胞遊走機構が全く不明のままである筋線維芽細胞において、細胞分化が進むことによってアクアポリンの発現量や局在が変化するのかどうかについて検討する(AQP1 発現が増加するとの予備知見がある)。また、細胞遊走刺激を与えた場合に、先導端にアクアポリンが局在するのか、アクチン重合に深く関わるコフィリンとともに検討する。
【項目6】カハール介在細胞の病態変化
カハール介在細胞は、消化管運動のペースメーカー機能を担う間葉系細胞である。炎症病態時に、カハール介在細胞のマーカーとされるc-kit 抗体の染色性が低下することはよく知られているが、細胞自体が消失するのか、あるいは脱分化するのかといった実態は解明されていない。本項目では、カハール介在細胞の傷害性/脱分化変化をマーカーとなり得る分子群を用いて解析する。また、カハール介在細胞の興奮性変化や、傷害後の回復過程についても明らかにする。
【項目7】PDGFRα陽性の線維芽細胞様細胞(新種の細胞)の病態変化
消化管筋層に存在する抑制性興奮機能を担う特殊な間葉系細胞であるが、研究分担者(飯野)により発見された新種の細胞である。病態時に筋線維芽細胞への分化が見られるのではないかと予想しているが、蛋白質レベルの変化に加え、生理機能としての興奮性変化を丹念調べて行く。一方、筋層におけるtenascin-C の恒常的な産生細胞ではないかとの予備知見もすでにある。
【項目8】ヒト臨床からのアプローチ
病態下におけるヒト臨床検体として、1) 小腸移植術後の移植腸管、2) 先天性の腸管運動障害を呈する小児のヒルシュスプルング病とその類縁疾患、を対象とする。小腸は体の外に面している臓器であることことから免疫機能が発達しているが、このことが原因となり、小腸移植は他臓器に比べて拒絶反応を起こしやすい。そして、慢性拒絶に至る過程において最大の特徴は線維化である。一方、ヒルシュスプルング病をはじめとする先天性腸管運動障害疾患においてカハール介在細胞の発現の減弱を認めることがあり、腸管運動 障害の一因になっていると考えられる。しかし、いずれも詳細な病態解明はされていない。本研究の基礎研究チームで得られた知見(主に分子情報)をヒト臨床サンプルに還元し、病態解明を図る。
平成26 年度以降の研究
26 年度以降も上記8項目の研究を継続実施する。予備知見も多いことから、計画通り進められると考えている。
表2 本研究で扱う間葉系細胞群(特徴と着眼点)
| (1)腸上皮細胞下の筋線維芽細胞 | 消化管粘膜の上皮直下には、収縮機能を持つ一層の星形の筋線維芽細胞が存在する。これらの細胞は細胞外基質を産生し、粘膜上皮細胞の分化増殖の足場となっている。粘膜直下というロケーションから上皮細胞との相互反応に注目したい。 |
|---|---|
| (2)肝星細胞(伊東細胞) 肝筋線維芽細胞 |
肝臓には類洞血管の周囲に存在する肝星細胞(伊東細胞)と門脈周囲に筋線維芽細胞が存在する。炎症にともなうこれら2 つの細胞の筋分化プロセス、特に転写調節系の差異を比較論的に解析する。 |
| (3)腸上皮細胞 | 腸上皮細胞としては、ラット小腸由来のIEC-6 を用いる。静止時には腸上皮としての性質をよく保持しており、上皮・間葉転換研究にも数多く使用されているが、収縮・運動機能という観点からの報告は少ない。 |
| (4)血管内皮細胞 | 炎症により組織に障害刺激が加わると、血管内皮細胞の活性型(分泌型)への変換、活性化内皮細胞からの液性因子の遊離、活性化内皮細胞を介した血液から組織へのマクロファージや各種白血球の浸潤・遊走が起こる。さらにTGFβにさらされると、内皮・間葉転換(Endo-MT)も起こる。一方、炎症部位における血管透過性亢進には収縮要素であるアクトミオシン系が関わっており、運動系という点では欠かせない細胞である。 |
| (5)中皮細胞 | 体腔内臓器表面とこれに連なる体腔内側の表面は中皮細胞と呼ばれる。病態としては、術後の臓器癒着や長期の腹膜透析に伴う腹膜炎や被嚢性腹膜硬化症の原因となる。収縮・運動機能という観点からの報告は皆無である。 |
| (6)カハール介在細胞 | 肥満細胞と同様にc-kit を発現する特異な間葉系細胞で、盛んに興奮性活動電位を発生し、心臓でいえば刺激伝導系に相当するペースメーカー細胞と考えられている。炎症時にはc-kit を失い変異するが、その詳細は未解明である。 |
| (7)PDGFRα陽性の線維芽細胞様細胞 | α型PDGF 受容体とSK3 チャネルを発現する線維芽細胞様の細胞である。ATPによって外向き電流が流れることから、カハール介在細胞とは逆の、抑制性の興奮細胞と考えられている。病態変化についての知見は皆無である。 |

