|
|---|
�ȑO����A���ǂ̔S�����ǂƒ����t���[���̗���͖��ڂɊ֘A���邱�Ƃ��m���Ă��܂����B��U�S���ɉ��ǂ��N����A���ꂪ�i�s���ĉ��ǂ��ؑw�ɋy�ԂƁA���ǂ̉^���@�\��Q�������N�����B���lj^���͒����t���[���ێ��̍ŏd�v�Ȉ��q�ł���A�����ɗ��ꂪ������ƔS�����ǂ��������A��̈��z�̏�Ԃ������炵�Ă��܂����ƂɂȂ�܂��B�������́A�q���V���X�v�����O�a���f�����b�g�ɂ����āA�����t���[���̗���ɕt�����ċؑw�ł݂��鉊�ǔ������A�ؑw�Ԑ_�o�p�����ɏ�݂���}�N���t�@�[�W�Ɉˑ�����̂ł͂Ȃ����Ƃ̉���̂��ƂɎ������s���A������ؖ����A����ɖ{�a�ԃ��f���ł݂���^���@�\��Q�Ɋւ�邱�Ƃ����o���܂����B�����ł́A�����t���[���ƒ��lj^���@�\��Q�Ƃ̊W���Љ�邱�Ƃɂ��܂��B(�����ۊw� 19, 9-16, 2005�����ς��Čf�ڂ��Ă��܂��BAm J Physiol Gastrointest Liver Physiol.287, G638-G646���x�[�X�ƂȂ��Ă��܂��B)
�͂��߂�
�����͗L�v�Ȓ����t���[���Ƌ������Đ����Ă��邪�A�����t���[���̐���p�^�[���́A�ۂ̕������x�Ə����ǂ̏㕔���牺���ւ̗���Ƃ����Q�̑傫�ȗv�f�̃o�����X�̏�ɐ��藧���Ă���B���̊e���ʂɂ͓K�ȍۂ��K���Ȑ��ő��݂��邱�Ƃ��d�v�ł����B���ǐ��������iIBD�j�̃��f�������ɂ������́A������̏ꍇ�����ۏ�Ԃł͒����͔������Ȃ��B�]���āA�����t���[���̑��݂����S�����ǂ̔��ǂɏd�v�ł���ƈ�ʂɔF������Ă���(1)�B�Ƃ���ŁA���ǂ̃o���A�[�@�\���l����ƁA�S�������ǂŔj��ꍇ�A�[�w�̋ؑw���ۂ̐N�����A��X�̓őf��ٕ��ɒ��ڂ��炳��邱�ƂɂȂ�B�����āA�L�Q�ȍۂ�ٕ���̊O�֔r�o����@�\�́A���ǂ̉^���@�\�Ɉˑ�����B���������āA�S�����ǂ��ؑw�ɂ܂ŋy�ыؑw�̉^���@�\����Q�����ƁA�����ǑS�̂���������Ɨ\�z�ł����B
�����t���[���ƒ��S���̊W�ɂ��Ă����͐��������邪�A�����t���[�����������^���Ƃ̊W�ׂ������͋ɂ߂ď��Ȃ��B����ɁA�ؑw�̉��ǂ��̂��̂̌������ɂ߂ĖR�����AIBD�ł����Δ�������^���@�\��Q�̋@���͂قƂ�ǖ��炩�ɂ���Ă͂��Ȃ��̂�����ł���B�{�e�ł́A�l�X�Ȓ������Ŗ��ƂȂ�����lj^���@�\��Q�Ƌؑw���ǔ����̊W�ɂ��Ă̍ŐV�̒m�����Љ�A�����lj^���@�\���R���g���[������v�f�Ƃ��Ă̒����t���[���A���邢�͒����t���[�����R���g���[������v�f�Ƃ��Ă̏����lj^���@�\�̈Ӌ`�ɂ��āA��X���ŋߍs�����q���V���X�v�����O�a�������ɍl���Ă݂����B
�P�D���lj^���͒����t���[���ɂ���ĉe������邩
�����t���[���́A�������̋ێ킪�K�Ȓ��ǂ̕��ʂɂ����đ��B�E�蒅���Ă��邱�Ƃ��d�v�ł���B���̒����t���[���̃v���t�B�[���́A�l�X�ȗv���i�Ⴆ�ApH��_�`�A�����y�f�Ȃǁj�ʼne�����邪�A�Ȃ��ł��������e���̗���A���Ȃ킿�����lj^���������t���[���v���t�B�[�����K�肷��d�v�ȗv�f�ƍl�����Ă���B
�����ŁA�����t���[�����^���@�\�ɉe����^���邱�Ƃ���������̘_�����Љ��B�Ⴆ�A���ۓ������ǁi�����j�ɂ��������e���A�����x�͐��퓮���ɔ�ׂĒx�����A�����ۑp��蒅������ƒZ�k����(2, 3)�B�܂��A���ۓ�����Lactobacillus acidophilus��Bifidobacterium bifidum�𓊗^����ƗA�����x�͒ʏ퓮���̑��x�ɋ߂Â����Ƃ��ώ@����Ă���(4)�B
 �@�@�}�P
�@�@�}�P
�����̉^���́A���ƐH����ł��ꂼ����L�̎��k�p�^�[���������B���ł͐Î~���i�T���j�ƕs�K���ȏ��U���̎��k�Q�i�U���j�ƁA����ɑ������p�x�̋K���I�ȑ�U���̎��k�Q�i�V���j�������B�����V���̎��k�Q�́A�����NJԌ��`�d�����kinterdigestive migrating contraction�iIMC�j�ƌĂ�A従��^���̎w�W�Ƃ����B������IMC���N�����d�C�����͓`�d���ؕ��d�Qmigrating myoelectric complex�iMMC�j�Ƃ��A従����ɂ��̊Ԋu���Z�k����A�����؊��������i����B���̂l�l�b�ɑ��āA�����t���[�����e����^����Ƃ���������B�Ⴆ�AMicrococcus luteus ��Escherichia coli�́AMMC�Ԋu���������A�����lj^�������コ����B���Ȃ킿�A���̂͋ۂ̎����قȐ�����F�m���A���炩�̋@������ĉ^���n���ԐړI�ɐ��䂵�Ă���Ƒz�������B���ہAClostridium difficile ���Y������Toxin A���Ain vitro�̏������ŏ����Ǖ����̕������_�o�����ɉe������Ƃ̕�����(5)�B����ɁA�R�ۖ�̌o�����^�������lj^���ɉe�����邱�Ƃ��m���Ă��邪�A����������t���[���̕ω����^���n�ɉe����^�������ʂƉ��߂����B
�Ƃ���ŁA�֔�ȂǏ����lj^�����}�����ꂽ��Ԃł́A���C���ۂ��������A�D�C���ۂƕ��s�ہiClostridium perfringens�Ȃǁj���������邱�Ƃ��m���������B����ɁA�����q�l���^�ɂ�������ǂɂ����ăR�����쓮���_�o������}�������lj^����}�����邱�Ƃɂ�������I�ɕ֔���N�����ƁA�����t���[���̒��ōD�C���ۂ̑̓��ڍs�p�x���������邱�Ƃ�����Ă���(6)�B���̗l�ɁA�����t���[���v���t�B�[�������lj^���ɉe����^���������A���ǂ̉^���������t���[���̃v���t�B�[���ɉe������Ƃ����\�}�������яオ���Ă��邪�A�����̌����ł͂��̋@���ɂ܂œ��ݍ���Ō�������Ă���킯�ł͂Ȃ��B
�Q�D���lj^���@�\��Q
�P�j���lj^���@�\�̗���
���̒f�Ђ�E�o���h�{�t�Ŗ��������V���[���ɒu���Ă��A���h��従��^�����ώ@���邱�Ƃ��ł��A����͐_�o������}������e�g���h�g�L�V���ŗ}�������B���̗l�Ȏ�������A�ÓT�I�ȃX�L�[���Ƃ��āA従��^���̂悤�ȃV�X�e�������ꂽ�����lj^���@�\�ɂ͋ؑw�Ԃɑ��݂���_�o�l�b�g���[�N���ւ��Ƃ���Ă����B�Ƃ��낪�A���f�Ђ���_�o�l�b�g���[�N���܂܂�Ȃ��悤�ȋؑg�D��Ђ��쐬���Ă��A�����I�Ȏ��k���J��Ԃ��B���̎��k�̓e�g���h�g�L�V���ŎՒf����Ȃ��̂ŁA�،����ł���ƍl�����Ă���B�������ƁA���̋،������k�̋@���͕s���ł��������A�ŋ߂��̖�肪�悤�₭�������ꂽ�B���̋@�\��S���̂��A�ؑw�Ԑ_�o�p�ɖ��ɕ��z����J�n�[����ݍזE�iInterstitial cell of Cajal : ICC�j�ł���(7-9)�BICC�͋ؑw�Ԑ_�o�p�����ł͂Ȃ������Njؑw�ɂ��L�����z����B�O�҂̓y�[�X���[�J�[�@�\���͂����A��҂͎h���`���@�\�����ƍl�����Ă���B�@�\�I�ɂ͐S�̎h���`���n�זE�Ƃ悭���Ă���ƌ�����BICC��c-kit����ٓI�ɔ�������̂ŁA�����؍זE�Ƃ͗e�Ղɋ�ʂ��邱�Ƃ��o����B�ŋ߁A�J�n�[����ݍזE���A�_�o�ƕ����̒��ԂɈʒu���Đ_�o�����̒���҂Ƃ��ċ@�\����\�����w�E����A����ɒ��ڂ���Ă���B
�Q�j�ؑw�̖Ɖu�n�F��^�}�N���t�@�[�W
���ǂ͔S������Đ��̊O�Ɛڂ��Ă���A��ɕa���̂̊�����ٕ��̐N���̊댯�ɂ��炳��Ă���B���ǔS���ɂ͐������̌ŗL�̃����p���ق��A�p�C�G����Ǘ������p���߂Ȃǂ̃����p�g�D�����݂��邪�A�����ɂ͑����̏�^�}�N���t�@�[�W�����݂��A�S���a���ɐ[���ւ���Ă����B���ǔS���Ɖu�́A�Ɖu�w�̒��ł���v�Ȍ����e�[�}�̈�ł���A����ɂ͖c��Ȍ���������B����A���ǐ��������iIBD�j�Ȃǂ̏d�x�ȔS�����ǂ������ɂ͉^���@�\��Q���������Ƃ��������A�^���@�\�̗���͒����t���[���ُ̈퉻�������炵�A���ǂ̈����ɑ傫����^���Ă���\�����l������B���Ȃ킿�A�S���A�ؑw�����Ē����t���[���̂R�҂݂͌��ɉe���������Ē��̃z���I�X�^�V�X���`�����Ă���ƍl�����邪�A�S���a�ς����ł͂Ȃ����̗l�ȕ��G�n�̔j�]�̉��Ǖa�Ԃƌ��邱�Ƃ��o����B
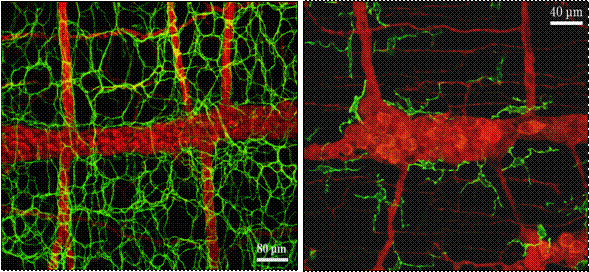
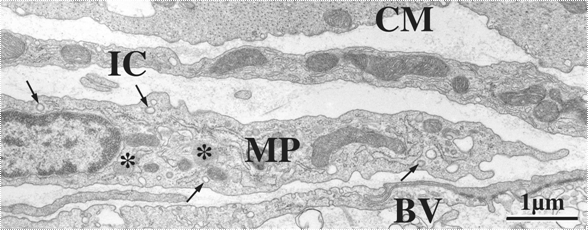
�Ƃ�����A�����ǂ̋ؑw���_�o�p�̕����ɂ́A�y�[�X���[�J�[�@�\���͂����ƍl�����Ă���ICC������ő��݂������A���͂���Ɠ����ʂɏ�^�}�N���t�@�[�W���K�����������z���Ă����i�}�Q�j�B�d�q�������ɂ��ώ@�ɂ��A�����Njؑw�̏�^�}�N���t�@�[�W�ׂ͍��Z���U���l�ˋN��L���A��r�I�d�q���x�̒Ⴂ�זE���������A�����̃��C�\�]�[����핢���E�icoated vesicle�j��L���邱�Ƃœ����Â����A���͂̕�����_�o�v�f�AICC���Ǝ��ʂ�����i�}�R�j�B�P���|�{�������т���A�@�\�I�ɂ��}�N���t�@�[�W�ł��邱�Ƃ��m�F����Ă���(10)�B
 �@�}�Q
�@�}�Q
 �@�@�}�R
�@�@�}�R
�����ŁA�ؑw�̏�^�}�N���t�@�[�W�̌����̗��j��U��Ԃ��Ă݂�ƁA����ȋؑg�D�Ƀ}�N���t�@�[�W�l�זE�����݂���Ƃ����L�ڂ�100�N�ȏ���O����Ȃ���Ă������A�L���F�m����邱�Ƃ͂Ȃ��A�܂����ۉ�זE��ICC�ƍ�������邱�Ƃ������ł������B1985�N��Mikkelsen���(11)�A��Ƃ��ăz�[���}�E���g�W�{�ŏڍׂɊώ@���A�ؑw�Ԑ_�o�p�Ȃ�тɟ����̏c���ؑw���ɏ�^�}�N���t�@�[�W���K���I�ɕ��z���邱�Ƃ�����B���̌��������������ƂȂ�A�ؑw����^�}�N���t�@�[�W�̋@�\�̉�͂��s����悤�ɂȂ����B�ؑw�Ԑ_�o�p�ɑ��݂����^�}�N���t�@�[�W�́A�_�o�ߍזE�Ƌߐڂ��Ă��邾���ł͂Ȃ��A���lj^���̃y�[�X���[�J�[�זE�Ƃ��Ēm����ICC�ɂ��ڂ��Ă���B���Ȃ킿�A��^�}�N���t�@�[�W��ICC���邢�͐_�o�Ƃ����������؉^���@�\�𐧌䂷��זE�n�Ƃ̃N���X�g�[�N����āA���lj^���@�\�̒��߂Ɋւ���Ă���\�����l�����A���ڂ���Ă���B
�R�j��^�}�N���t�@�[�W�ƒ��lj^���@�\�Ɋւ��錤��
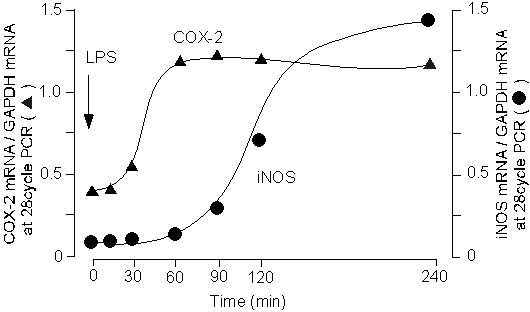
1985�N��Mikkelsen��̕�������A���̍זE���ؑw�̉��ǂƂ���ɑ����^���@�\�̏�Q�Ɋւ��ł��낤���Ƃ͗e�Ղɑz������Ă������A�@�\�ʂł̌����͐i�܂Ȃ������BEskandari���(12)�A�s���ǎ��ɐ���������lj^���@�\��Q�ɂ��Ă̌������s���ALPS�����b�g�ɒ��˂����ۂɁA���Njؑw�̈�_�����f�iNO�j�Y�����������邱�ƁA�����Ă���NO���ؑw�ԏ�^�}�N���t�@�[�W�ɗU��������U���^NO�����y�f�iiNOS�j�ɋN�����邱�Ƃ����߂ĕ����B
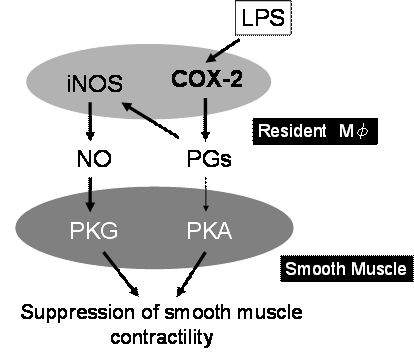
��X�́A���̏�^�}�N���t�@�[�W�̊������̕��q�@���𖾂炩�ɂ��邽�߁Ain vitro�̎����n�Ō�����������(13, 14)�B���̌��ʁA�����Ǒg�D��LPS�𓊗^����Ƌؑw�ԏ�^�}�N���t�@�[�W��COX-2���U������ăv���X�^�O�����W�����Y������A�����v���X�^�O�����W���Y�����������ƂȂ�iNOS���U������邱�Ƃ��������ꂽ(14)�B����ALPS��in vitro�Ő����ԏ��u���������Ǒg�D�̎��k�@�\�͋����}�������B���k�ቺ�̌����𖾂炩�ɂ��邽�߂�NOS�j�Q��L-NMMA�𓊗^�����Ƃ�����k����50%���ACOX�j�Q�܃C���h���^�V���𓊗^�ɂ���قڊ��S�ɉ����B�����̐��т���ALPS�ɂ������ǎ��k�̗}���͏�^�}�N���t�@�[�W����COX-2�̗U���A�v���X�^�O�����W���Y���^�����؎��k�}���Ƃ����o�H�ƁA�v���X�^�O�����W�����I�[�g�N���C���ɓ����ď�^�}�N���t�@�[�W����iNOS�U���ANO�Y���ANO�ɂ�镽���؎��k�}���Ƃ����o�H�̂Q��ނ����邱�Ƃ��������ꂽ�B�ȏ�̐��т́A�����lj��ǂ���^�}�N���t�@�[�W����ď����lj^���@�\�ɑ傫�ȉe�����y�ڂ����Ƃ��������������A��������ƂȂ镪�q��COX-2�Ƃ������ƂɂȂ��i�}�S�A�T�j�B
 �@�@�}�S
�@�@�}�S
 �@�@�}�T
�@�@�}�T
����A���퓮���ł̋ؑw����^�}�N���t�@�[�W�ł́AMHC class II�R�������Ă���זE�͏��Ȃ��ALPS�łS�`�W���Ԏh������Ɣ����זE�̐�����������B���������āA�Î~���ɂ����Ă͋ؑw����^�}�N���t�@�[�W�̑啔���͕s�����̏�Ԃő��݂��Ă�����̂Ǝv����(13)�BTurler���(15)�ALPS�𓊗^�������b�g����E�o�������ǂ̋ؑw����^�}�N���t�@�[�W�ŁA�P���̗V�����q�ł���monocyte chemoattractant protein-1�iMCP-1�j�̔������㏸���邱�Ƃ��ώ@���Ă���B���̂��Ƃ́A���펞�ɂ͕s�����ł����^�}�N���t�@�[�W���A�������ǎ��ɂ�LPS�Ȃǂ̋ۑ̐����ɂ��h����[���Ƃ��Ċ���������邱�ƁA�����āA�}�N���t�@�[�W�̊��������N�_�ɁA�����̖Ɖu�n�זE���Q�����ċؑw���ǂǂ����邱�Ƃ���������B
�R�D�q���V���X�v�����O�a�ԃ��f�������ɂ�����ؑw�̉���
�P�j���_�o�ߒ��ǃ��b�g
���_�o�ߒ��ǃ��b�gAganglionosis Rat�iAR�j�̓E�C�X�^�[�������b�g�Ɩ�l�̌�z�ɂ�蓾��ꂽ�ˑR�ψّ̂ŁA�������邢�͉������璼���ɂ����Ē��ǐ_�o�߂��V�I�Ɍ������Ă���B���̕��ʂ�������N�����A����������ŐH��ɂ�鋐��ǂ܂��͋��匋���ǂǂ��A����1�����ȓ��Ɏ��S����(16, 17)�B���̂���AR�́A��V�I�Ȓ��ǐ_�o�ߌ��@�ɂ�鋐�匋���ǂ������N�����q���V���X�v�����O�a�̕a�ԃ��f���Ƃ��čl�����Ă���B�܂��AAR�̌�����`�q (sl) �͏���F�̒P��̈�`�l�����Ƃ�AAR�͗z���̂ł���A�����j���זE���������߁A�����ɂ킸���̗L�F��т̕������F�߂��邾���őS�g���F��тł���B����A����z���̂���уw�e���͓̂����S�̂���ю�ɔw���𒆐S�Ƃ����̂̈ꕔ�����L�F��тŕ����Ă���(16)�i�}�U���j�B���̌�AAR�ɂ����ăG���h�Z����B��e�̈�`�q�ɑ�1�G�L�\�������1�C���g�����ɂ܂�����301 bp �̌���������A���ꂪ���ǐ_�o�߂̌����ɂ�鋐��ǂ܂��͋��匋���ǁA����ёS�g���F�̌����ł��邱�Ƃ����炩�ɂȂ���(18)�i�}�U�E�j�B����ɁA���ǐ_�o�߂ƃ����j���זE�͓����_�o��זE���甭�����A�G���h�Z����B��e�̂���� endothelin-3 �̃V�O�i���`�B�n���A���̐_�o��זE�̔��B�ɕs���ł��邱�Ƃ����炩�ɂȂ���(19)�B
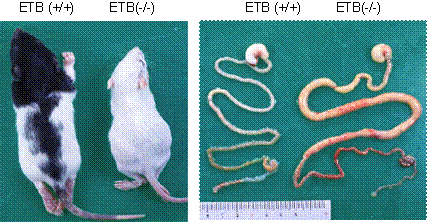 �@�@�}�U
�@�@�}�U
�Q�jAR�̏����c�啔�̒����t���[���Ƌؑw����
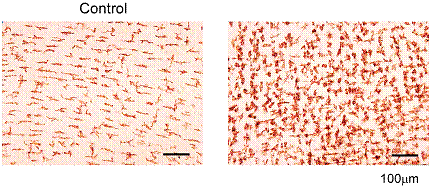
���S���̉��ǂɂ͒����t���[���ُ̈킪�[���ւ�邱�Ƃ���A�܂��n�߂�AR�̏����i�_�o�p������ȕ����j�̒����t���[������͂����B���Nj���іc���������AR�ł́A���ǖc�啔�i�j�ɂ����āA���ې��������������ɒʏ�͖Ӓ��Ō�����悤��Clostridia�����o�����A�܂�Streptococci�����������Ă���A�����t���[�����v���t�B�[�����傫���ω������ُ�Ȋ��ƂȂ��Ă����i�\�P�j�B����ɁA�S���ɂ����鋭�����Ǒ����m�F�����ƂƂ��ɁA�S�����̋ؑw�ɂ����Ă����ǍזE�̐Z��������ꉊ�ǔ������i��ł��邱�Ƃ��m�F���ꂽ�B�̓��ڍs���N�����L�Q�ȍۗނ̑����������ŁA���ʂ̍ۂ��S����ؑw�ɐN�����A���s����đS�g�ɓ`�d���Ă���\�����l����ꂽ�B
Bacterial groups Control ETB (-/-) Enterobacteriaceae 4.0 �} 1.2 9.1 �} 1.8** Enterococci 6.6 �} 1.1 9.7 �} 0.6** Staphylococci 5.4 �} 0.9 5.2 �} 1.1 Gram (-) aerobic rods 5.0 N.D. Yeasts N.D. 3.0 Lactobacilli 8.8 �} 0.3 9.1 �} 0.7 Bacteroidaceae 5.8 �} 1.3 10.0 �} 0.6** Bifidobacteria 5.0 N.D. Eubacteria 6.8 �} 2.1 9.9 �} 0.6** Clostridia N.D. 8.8 �} 1.1 Total count 8.9 �} 0.3 10.6 �} 0.5
�@�@�@�@�@�@�@�\�P
�@
����LPS�Z�x�𑪒肷��ƁA�L�ӂȑ������F�߂�ꂽ�B�����t���[�����傫���ω��������ǖc�啔�ł�LPS�Y�����l������ƁA�����ǕNjǏ��ɂ�����}�N���t�@�[�W���A�����Z�x���������Z�x��LPS�ɔ��I���ꊈ��������Ă���\�����z�肳���B���ɁAAR�̒��ǂ̋ؑw��E�o���A�T�C�g�J�C����mRNA�ʂ𑪒肵���Ƃ���AIL-1b��IL-6�̗L�ӂȑ������F�߂�ꂽ�i�}�V�j�B�܂����̎��AED2�R�̗z���̋ؑw����^�}�N���t�@�[�W�̐����Q�`�R�{�ɑ������Ă����i�}�W�j�B
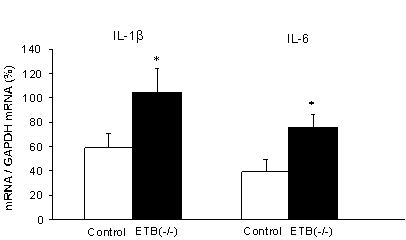 �@�@�}�V
�@�@�}�V
 �@�}�W
�@�}�W
�R�j���_�o�ߕ��ł͂Ȃ��c�啔�̉^���@�\��Q
����ɁAAR�̕Ǔ��_�o�p��L����㕔�̏����ǂ���֏��E�o���A���k�������ώ@����ƁA���Y���̗���Ƃ��������I�ȕω����F�߂�ꂽ�i�}�X�j�B�܂��A���������k�̕p�x���L�ӂɒቺ���Ă������A���ǂ��݂��Ȃ��㕔�̏\��w���ł͂��̗l�ȕω��͊ώ@����Ȃ������B����ɁAAR��ICC��c-kit�R�̂Ő��߂Ă݂�ƁA�ΏƂɔ�ׂ��̐����L�ӂɌ������Ă���A�`�ԕω����F�߂�ꂽ�i�}�P�O�j�B���Ȃ킿�A���lj^���̌����� ICC�̌����Ƃ��̃l�b�g���[�N�\���̕ω��ɂ����̂ł���ƍl����ꂽ(20)�B
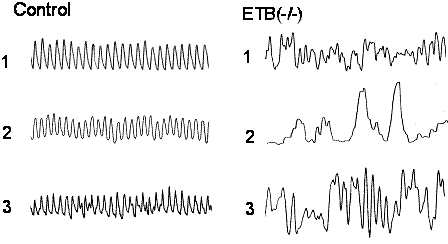 �@�@�}�X
�@�@�}�X
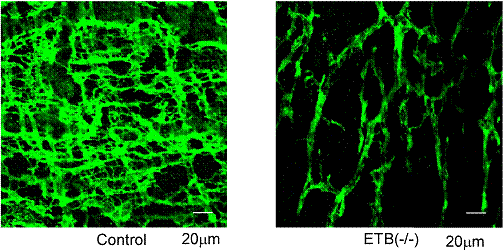 �@�@�}�P�O
�@�@�}�P�O
�S�j�q���V���X�v�����O�a���Nj���̂R�̌���
����܂ŁA�q���V���X�v�����O�a���Nj���̌����́A����従��̓����@�\�������ǂ�_�o�T�[�L�b�g�̌����ɂ��ƍl�����Ă����B�������A�{�����ɂ��q���V���X�v�����O�a�̐i�W�ɂ��L�_�o�ߕ��ɂ����ǂ��N����A�����ɉ^���@�\��Q�������ď㕔����̋�o�͂����ނ��邱�Ƃ�����ƂȂ邱�Ƃ������ꂽ�B�d�x�̃q���V���X�v�����O�a�ł͖��_�o�ߒ��ǂ̐؏��p���{����邪�A����ł��Ȍ�֔̕�X�������S�ɉ��P�����킯�ł͂Ȃ��A�ۑ��I�ȗÖ@�������B���̎��A���lj��ǂ�@���ɃR���g���[�����邩�A���Ȃ킿�����t���[���̃R���g���[���̏d�v�������������B
�Ƃ���ŁAAR�̖��_�o�ߕ����̒��ǂ̕����ؑw�͑��B���Ĕ�����A�܂������؎��g�̎��k�������i���A���̂��Ƃ�����̂����P�̗v���Ƃ��Ȃ��Ă���(21)�B���̗l�ɁA�q���V���X�v�����O�a���Nj���̌����́A�_�o�߂��������Ă��邱�Ƃɉ����A���Ȃ��Ƃ��Q�̓I�v�����֗^���Ă�����̂ƍl������(20, 21)�B
�S�D�܂Ƃ�
�@���ǂ��������ł͔S���̉��ǂ����ƂȂ邪�A�����ɏ����ǂ̉^���@�\����Q����邱�Ƃɂ����ڂ��Ȃ���Ȃ�Ȃ��B���Ȃ킿�A�S���̉��ǂ��i�s����ƁA���̉��ǔ����͎���ɋؑw�ւ��`���B�ؑw�ɉ��ǂ��y�ԂƐ_�o�EICC�E�����ɂ���č\�z�����^���n����Q�����B���̎��A�����t���[���̕ω����Ɖu��������ďd�v�Ȗ������ʂ����ƍl������B���Ȃ킿�A�^���@�\��Q�ɂ���Ē����t���[���͗���A���̌��ʊǍo���ɗl�X�ȗL�Q�������~�ς���A�ۂ̑̓��ڍs���i�s���A�S���̉��ǂ���������B�S�����ǂ̈����͂���ɋؑw���ǂ����������A���z�ɓ�����̂Ƒz�������B���̎��d�v�Ȗ�����S���̂��A�ؑw�̎��R�Ɖu�n�������ǂ�ƍl�������^�}�N���t�@�[�W�ł���B
1. Rennick, D.M., and Fort, M.M. 2000. Lessons from genetically engineered animal models. XII. IL-10-deficient (IL-10(-/-) mice and intestinal inflammation. Am J Physiol Gastrointest Liver Physiol 278:G829-833.
2. Caenepeel, P., Janssens, J., Vantrappen, G., Eyssen, H., and Coremans, G. 1989. Interdigestive myoelectric complex in germ-free rats. Dig Dis Sci 34:1180-1184.
3. Husebye, E., Hellstrom, P.M., and Midtvedt, T. 1992. Introduction of conventional microbial flora to germfree rats increases the frequency of migrating myoelectric complex. J. Gastrointest. Mot. 4:39-45.
4. Husebye, E., Hellstrom, P.M., Sundler, F., Chen, J., and Midtvedt, T. 2001. Influence of microbial species on small intestinal myoelectric activity and transit in germ-free rats. Am J Physiol Gastrointest Liver Physiol 280:G368-380.
5. Xia, Y., Hu, H.Z., Liu, S., Pothoulakis, C., and Wood, J.D. 2000. Clostridium difficile toxin A excites enteric neurones and suppresses sympathetic neurotransmission in the guinea pig. Gut 46:481-486.
6. Nieuwenhuijs, V.B., van Duijvenbode-Beumer, H., Verheem, A., Visser, M.R., Verhoef, J., Gooszen, H.G., and Akkermans, L.M. 1999. The effects of ABT-229 and octreotide on interdigestive small bowel motility, bacterial overgrowth and bacterial translocation in rats. Eur J Clin Invest 29:33-40.
7. Sanders, K.M., Ordog, T., and Ward, S.M. 2002. Physiology and pathophysiology of the interstitial cells of Cajal: from bench to bedside. IV. Genetic and animal models of GI motility disorders caused by loss of interstitial cells of Cajal. Am J Physiol Gastrointest Liver Physiol 282:G747-756.
8. Sanders, K.M., Ordog, T., Koh, S.D., Torihashi, S., and Ward, S.M. 1999. Development and plasticity of interstitial cells of Cajal. Neurogastroenterol Motil 11:311-338.
9. Sanders, K.M. 1996. A case for interstitial cells of Cajal as pacemakers and mediators of neurotransmission in the gastrointestinal tract. Gastroenterology 111:492-515.
10. Ozaki, H., Kawai, T., Shuttleworth, C.W., Won, K.J., Suzuki, T., Sato, K., Horiguchi, H., Hori, M., Karaki, H., Torihashi, S., et al. 2004. Isolation and characterization of resident macrophages from the smooth muscle layers of murine small intestine. Neurogastroenterol Motil 16:39-51.
11. Mikkelsen, H.B., Thuneberg, L., Rumessen, J.J., and Thorball, N. 1985. Macrophage-like cells in the muscularis externa of mouse small intestine. Anat Rec 213:77-86.
12. Eskandari, M.K., Kalff, J.C., Billiar, T.R., Lee, K.K., and Bauer, A.J. 1999. LPS-induced muscularis macrophage nitric oxide suppresses rat jejunal circular muscle activity. Am J Physiol 277:G478-486.
13. Torihashi, S., Ozaki, H., Hori, M., Kita, M., Ohota, S., and Karaki, H. 2000. Resident macrophages activated by lipopolysaccharide suppress muscle tension and initiate inflammatory response in the gastrointestinal muscle layer. Histochem Cell Biol 113:73-80.
14. Hori, M., Kita, M., Torihashi, S., Miyamoto, S., Won, K.J., Sato, K., Ozaki, H., and Karaki, H. 2001. Upregulation of iNOS by COX-2 in muscularis resident macrophage of rat intestine stimulated with LPS. Am J Physiol Gastrointest Liver Physiol 280:G930-938.
15. Turler, A., Schwarz, N.T., Turler, E., Kalff, J.C., and Bauer, A.J. 2002. MCP-1 causes leukocyte recruitment and subsequently endotoxemic ileus in rat. Am J Physiol Gastrointest Liver Physiol 282:G145-155.
16. Ikadai H, F.H., Agematsu Y, et al. 1979. Observation of congenital aganglionosis rat(Hirschsprung's disease rat) and its genetical analysis. Cong Anom:31-36.
17. Ikadai H, S.K., Fujita H, et al. 1981. Animal model of human disease: Hirschsprung's disease. Comput Pathol Bull. 13:3-4.
18. Ceccherini, I., Zhang, A.L., Matera, I., Yang, G., Devoto, M., Romeo, G., and Cass, D.T. 1995. Interstitial deletion of the endothelin-B receptor gene in the spotting lethal (sl) rat. Hum Mol Genet 4:2089-2096.
19. Hosoda, K., Hammer, R.E., Richardson, J.A., Baynash, A.G., Cheung, J.C., Giaid, A., and Yanagisawa, M. 1994. Targeted and natural (piebald-lethal) mutations of endothelin-B receptor gene produce megacolon associated with spotted coat color in mice. Cell 79:1267-1276.
20. Suzuki, T., Won, K.J., Horiguchi, K., Kinoshita, K., Hori, M., Torihashi, S., Momotani, E., Itoh, K., Hirayama, K., Ward, S.M., et al. 2004. Muscularis inflammation and the loss of interstitial cells of Cajal in the endothelin ETB receptor null rat. Am J Physiol Gastrointest Liver Physiol.287, G638-G646.
21. Won, K.J., Torihashi, S., Mitsui-Saito, M., Hori, M., Sato, K., Suzuki, T., Ozaki, H., and Karaki, H. 2002. Increased smooth muscle contractility of intestine in the genetic null of the endothelin ETB receptor: a rat model for long segment Hirschsprung's disease. Gut 50:355-360.