






しっかり学習:消化管運動機能障害と腸内フローラ/筋層免疫系
以前から、腸管の粘膜炎症と腸内フローラの乱れは密接に関連することが知られてきました。一旦粘膜に炎症が起こり、これが進行して炎症が筋層に及ぶと、腸管の運動機能障害を引き起こす。腸管運動は腸内フローラ維持の最重要な因子であり、ここに乱れが生じると粘膜炎症が悪化し、一つの悪循環の状態をもたらしてしまうことになります。私たちは、ヒルシュスプルング病モデルラットにおいて、腸内フローラの乱れに付随して筋層でみられる炎症反応が、筋層間神経叢部分に常在するマクロファージに依存するのではないかとの仮定のもとに実験を行い、これを証明し、さらに本病態モデルでみられる運動機能障害に関わることを見出しました。ここでは、腸内フローラと腸管運動機能障害との関係を紹介することにします。(腸内細菌学会誌 19, 9-16, 2005を改変して掲載しています。Am J Physiol Gastrointest Liver Physiol.287, G638-G646がベースとなっています。)
はじめに
動物は有益な腸内フローラと共存して生きているが、腸内フローラの数やパターンは、細菌の分裂速度と消化管の上部から下部への流れという2つの大きな要素のバランスの上に成り立っている。腸の各部位には適切な細菌が適当な数で存在することが重要でありる。炎症性腸疾患(IBD)のモデル動物においては、いずれの場合も無菌状態では腸炎は発生しない。従って、腸内フローラの存在が腸粘膜炎症の発症に重要であると一般に認識されている(1)。ところで、腸管のバリアー機能を考えると、粘膜が炎症で破壊され場合、深層の筋層も細菌の侵入を受け、種々の毒素や異物に直接さらされることになる。そして、有害な細菌や異物を体外へ排出する機能は、腸管の運動機能に依存する。したがって、粘膜炎症が筋層にまで及び筋層の運動機能が障害されると、腸炎症全体が悪化すると予想できる。
腸内フローラと腸粘膜の関係についての報告は数多くあるが、腸内フローラと消化管運動との関係を調べた研究は極めて少ない。さらに、筋層の炎症そのものの研究も極めて乏しく、IBDでしばしば発生する運動機能障害の機序はほとんど明らかにされてはいないのが現状である。本稿では、様々な腸疾患で問題となる消化管運動機能障害と筋層炎症反応の関係についての最新の知見を紹介し、消化管運動機能をコントロールする要素としての腸内フローラ、あるいは腸内フローラをコントロールする要素としての消化管運動機能の意義について、我々が最近行ったヒルシュスプルング病研究を例に考えてみたい。
1.腸管運動は腸内フローラによって影響されるか
腸内フローラは、ある特定の菌種が適切な腸管の部位において増殖・定着していることが重要である。この腸内フローラのプロフィールは、様々な要因(例えば、pHや胆汁、消化酵素など)で影響を受けるが、なかでも腸管内容物の流れ、すなわち消化管運動が腸内フローラプロフィールを規定する重要な要素と考えられている。
ここで、腸内フローラが運動機能に影響を与えることを示す幾つかの論文を紹介する。例えば、無菌動物腸管(小腸)における内容物輸送速度は正常動物に比べて遅いが、正常細菌叢を定着させると短縮する(2, 3)。また、無菌動物にLactobacillus acidophilusとBifidobacterium bifidumを投与すると輸送速度は通常動物の速度に近づくことも観察されている(4)。
 図1
図1
小腸の運動は、空腹期と食後期でそれぞれ特有の収縮パターンを示す。空腹期では静止期(Ⅰ相)と不規則な小振幅の収縮群(Ⅱ相)と、これに続く高頻度の規則的な大振幅の収縮群(Ⅲ相)が現れる。このⅢ相の収縮群は、消化管間欠伝播性収縮interdigestive migrating contraction(IMC)と呼ばれ、蠕動運動の指標とされる。そしてIMCを起こす電気活動は伝播性筋放電群migrating myoelectric complex(MMC)とよばれ、蠕動時にその間隔が短縮され、平滑筋活動が亢進する。このMMCに対して、腸内フローラが影響を与えるという報告がある。例えば、Micrococcus luteus とEscherichia coliは、MMC間隔を延長し、消化管運動を減弱させる。すなわち、生体は菌の持つ特異な成分を認知し、何らかの機序を介して運動系を間接的に制御していると想像される。実際、Clostridium difficile が産生するToxin Aが、in vitroの条件下で消化管平滑筋の副交感神経活動に影響するとの報告もある(5)。さらに、抗菌薬の経口投与が消化管運動に影響することも知られているが、これも腸内フローラの変化が運動系に影響を与えた結果と解釈される。
ところで、便秘など消化管運動が抑制された状態では、嫌気性菌が減少し、好気性菌と腐敗菌(Clostridium perfringensなど)が増加することが知られている。さらに、モルヒネ投与により消化管においてコリン作動性神経活動を抑制し腸管運動を抑制することにより実験的に便秘を起こすと、腸内フローラの中で好気性菌の体内移行頻度が増加することも報告されている(6)。この様に、腸内フローラプロフィールが腸管運動に影響を与える一方で、腸管の運動が腸内フローラのプロフィールに影響するという構図が浮かび上がってくるが、多くの研究ではその機序にまで踏み込んで研究されているわけではない。
2.腸管運動機能障害
1)腸管運動機能の理解
腸の断片を摘出し栄養液で満たしたシャーレに置いても、立派な蠕動運動を観察することができ、これは神経興奮を抑制するテトロドトキシンで抑制される。この様な事実から、古典的なスキームとして、蠕動運動のようなシステム化された消化管運動機能には筋層間に存在する神経ネットワークが関わるとされてきた。ところが、腸断片から神経ネットワークが含まれないような筋組織条片を作成しても、自発的な収縮を繰り返す。この収縮はテトロドトキシンで遮断されないので、筋原性であると考えられている。長いこと、この筋原性収縮の機序は不明であったが、最近この問題がようやく解決された。その機能を担うのが、筋層間神経叢に密に分布するカハール介在細胞(Interstitial cell of Cajal : ICC)である(7-9)。ICCは筋層間神経叢だけではなく消化管筋層にも広く分布する。前者はペースメーカー機能をはたし、後者は刺激伝導機能を持つと考えられている。機能的には心筋の刺激伝導系細胞とよく似ていると言える。ICCはc-kitを特異的に発現するので、平滑筋細胞とは容易に区別することが出来る。最近、カハール介在細胞が、神経と平滑筋の中間に位置して神経興奮の仲介者として機能する可能性が指摘され、さらに注目されている。
2)筋層の免疫系:常在型マクロファージ
腸管は粘膜を介して生体外と接しており、常に病原体の感染や異物の侵入の危険にさらされている。腸管粘膜には数多くの固有のリンパ球ほか、パイエル板や孤立リンパ小節などのリンパ組織が存在するが、ここには多数の常在型マクロファージが存在し、粘膜病変に深く関わっている。腸管粘膜免疫は、免疫学の中でも主要な研究テーマの一つであり、これには膨大な研究がある。一方、炎症性腸疾患(IBD)などの重度な粘膜炎症を伴う腸炎には運動機能障害が伴うことが多いが、運動機能の乱れは腸内フローラの異常化をもたらし、炎症の悪化に大きく寄与している可能性が考えられる。すなわち、粘膜、筋層そして腸内フローラの3者は互いに影響しあって腸のホメオスタシスを形成していると考えられるが、粘膜病変だけではなくこの様な複雑系の破綻を腸の炎症病態と見ることが出来る。
ところで、消化管の筋層間神経叢の平面には、ペースメーカー機能をはたすと考えられているICCが並んで存在するが、実はこれと同じ面に常在型マクロファージが規則正しく分布している(図2)。電子顕微鏡による観察により、消化管筋層の常在型マクロファージは細く短い偽足様突起を有し、比較的電子密度の低い細胞質に一次、二次のライソゾームや被覆小胞(coated vesicle)を有することで特徴づけられ、周囲の平滑筋や神経要素、ICC等と識別される(図3)。単離培養した成績から、機能的にもマクロファージであることが確認されている(10)。
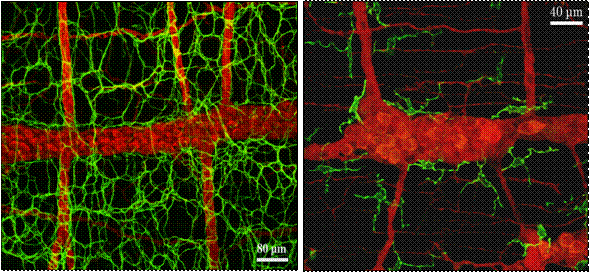 図2
図2
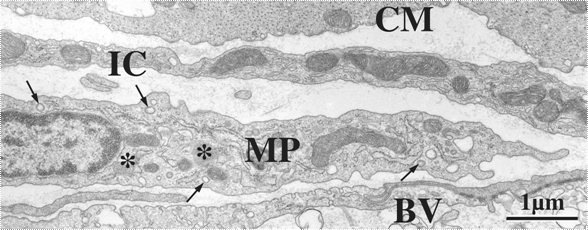 図3
図3
ここで、筋層の常在型マクロファージの研究の歴史を振り返ってみると、正常な筋組織にマクロファージ様細胞が存在するという記載は100年以上も前からなされていたが、広く認知されることはなく、また線維芽細胞やICCと混同されることがしばしばであった。1985年にMikkelsenらは(11)、主としてホールマウント標本で詳細に観察し、筋層間神経叢ならびに漿膜の縦走筋層側に常在型マクロファージが規則的に分布することを報告した。この研究がきっかけとなり、筋層内常在型マクロファージの機能の解析が行われるようになった。筋層間神経叢に存在する常在型マクロファージは、神経節細胞と近接しているだけではなく、腸管運動のペースメーカー細胞として知られるICCにも接している。すなわち、常在型マクロファージはICCあるいは神経といった平滑筋運動機能を制御する細胞系とのクロストークを介して、腸管運動機能の調節に関わっている可能性が考えられ、注目されている。
3)常在型マクロファージと腸管運動機能に関する研究
1985年のMikkelsenらの報告当初から、この細胞が筋層の炎症とこれに続く運動機能の障害に関わるであろうことは容易に想像されていたが、機能面での研究は進まなかった。Eskandariらは(12)、敗血症時に生じる消化管運動機能障害についての検討を行い、LPSをラットに注射した際に、腸管筋層の一酸化窒素(NO)産生が増加すること、そしてこのNOが筋層間常在型マクロファージに誘導される誘導型NO合成酵素(iNOS)に起因することを初めて報告した。
我々は、この常在型マクロファージの活性化の分子機序を明らかにするため、in vitroの実験系で検討を加えた(13, 14)。その結果、消化管組織にLPSを投与すると筋層間常在型マクロファージにCOX-2が誘導されてプロスタグランジンが産生され、このプロスタグランジン産生が引き金となりiNOSが誘導されることが示唆された(14)。一方、LPSをin vitroで数時間処置した消化管組織の収縮機能は強く抑制される。収縮低下の原因を明らかにするためにNOS阻害剤L-NMMAを投与したところ収縮は約50%回復し、COX阻害剤インドメタシンを投与によりほぼ完全に回復した。これらの成績から、LPSによる消化管収縮の抑制は常在型マクロファージ内のCOX-2の誘導、プロスタグランジン産生/平滑筋収縮抑制という経路と、プロスタグランジンがオートクラインに働いて常在型マクロファージ内のiNOS誘導、NO産生、NOによる平滑筋収縮抑制という経路の2種類があることが示唆された。以上の成績は、消化管炎症が常在型マクロファージを介して消化管運動機能に大きな影響を及ぼすことを示唆しており、その主役となる分子がCOX-2ということになる(図4、5)。
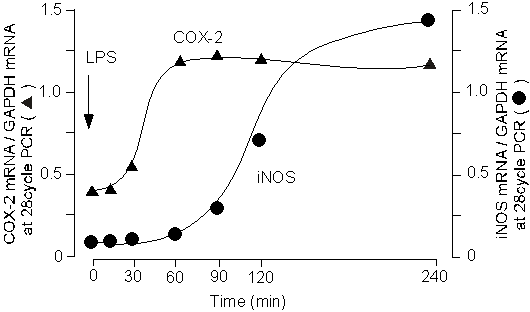 図4
図4
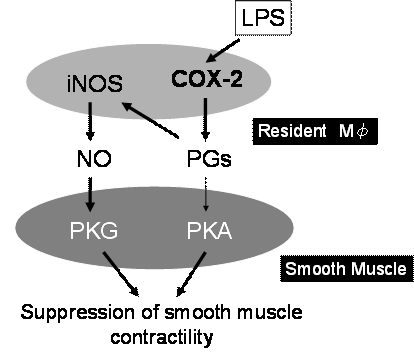 図5
図5
一方、正常動物での筋層内常在型マクロファージでは、MHC class II抗原を発現している細胞は少なく、LPSで4~8時間刺激すると発現細胞の数が増加する。したがって、静止時においては筋層内常在型マクロファージの大部分は不活性の状態で存在しているものと思われる(13)。Turlerらは(15)、LPSを投与したラットから摘出した腸管の筋層内常在型マクロファージで、単球の遊走因子であるmonocyte chemoattractant protein-1(MCP-1)の発現が上昇することを観察している。このことは、正常時には不活性である常在型マクロファージが、腸炎発症時にはLPSなどの菌体成分による刺激を端緒として活性化されること、そして、マクロファージの活性化を起点に、多くの免疫系細胞が参加して筋層炎症を発症させることを示唆する。
3.ヒルシュスプルング病態モデル動物における筋層の炎症
1)無神経節腸管ラット
無神経節腸管ラットAganglionosis Rat(AR)はウイスター今道ラットと野鼠の交配により得られた突然変異体で、回腸中部あるいは下部から直腸にかけて腸管神経節を先天的に欠損している。この部位が狭窄を起こし、それより口側で食塊による巨大回腸症または巨大結腸症を発症し、生後1ヶ月以内に死亡する(16, 17)。そのためARは、先天的な腸管神経節欠如による巨大結腸症を引き起こすヒルシュスプルング病の病態モデルとして考えられている。また、ARの原因遺伝子 (sl) は常染色体単一劣性の遺伝様式をとり、ARは劣性ホモ個体であり、メラニン細胞を欠くため、頭部にわずかの有色被毛の部分が認められるだけで全身白色被毛である。一方、正常ホモ個体およびヘテロ個体は頭部全体および主に背側を中心とした体の一部分が有色被毛で覆われている(16)(図6左)。その後、ARにおいてエンドセリンB受容体遺伝子に第1エキソンから第1イントロンにまたがる301 bp の欠失があり、これが腸管神経節の欠損による巨大回腸症または巨大結腸症、および全身白色の原因であることが明らかになった(18)(図6右)。さらに、腸管神経節とメラニン細胞は同じ神経堤細胞から発生し、エンドセリンB受容体を介した endothelin-3 のシグナル伝達系が、この神経堤細胞の発達に不可欠であることが明らかになった(19)。
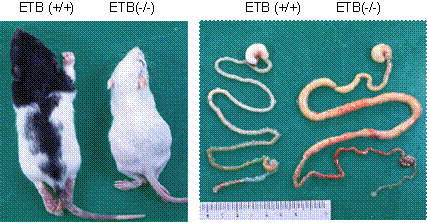 図6
図6
2)ARの小腸膨大部の腸内フローラと筋層炎症
腸粘膜の炎症には腸内フローラの異常が深く関わることから、まず始めにARの小腸(神経叢が正常な部分)の腸内フローラを解析した。腸管狭窄および膨大をおこすARでは、腸管膨大部(回腸)において、総細菌数が増加し、さらに通常は盲腸で見られるようなClostridia属が出現し、またStreptococci属が増加しており、腸内フローラのプロフィールが大きく変化した異常な環境となっていた(表1)。さらに、粘膜における強い炎症像が確認されるとともに、粘膜下の筋層においても炎症細胞の浸潤が見られ炎症反応が進んでいることが確認された。体内移行を起こす有害な細菌類の増加も顕著で、多量の細菌が粘膜や筋層に侵入し、血行を介して全身に伝播している可能性が考えられた。
Bacterial groups Control ETB (-/-) Enterobacteriaceae 4.0 ± 1.2 9.1 ± 1.8** Enterococci 6.6 ± 1.1 9.7 ± 0.6** Staphylococci 5.4 ± 0.9 5.2 ± 1.1 Gram (-) aerobic rods 5.0 N.D. Yeasts N.D. 3.0 Lactobacilli 8.8 ± 0.3 9.1 ± 0.7 Bacteroidaceae 5.8 ± 1.3 10.0 ± 0.6** Bifidobacteria 5.0 N.D. Eubacteria 6.8 ± 2.1 9.9 ± 0.6** Clostridia N.D. 8.8 ± 1.1 Total count 8.9 ± 0.3 10.6 ± 0.5
表1
血中LPS濃度を測定すると、有意な増加が認められた。腸内フローラが大きく変化した腸管膨大部でのLPS産生を考慮すると、消化管壁局所におけるマクロファージが、血中濃度よりも高い濃度のLPSに曝露され活性化されている可能性が想定される。次に、ARの腸管の筋層を摘出し、サイトカインのmRNA量を測定したところ、IL-1bとIL-6の有意な増加が認められた(図7)。またこの時、ED2抗体陽性の筋層内常在型マクロファージの数が2~3倍に増加していた(図8)。
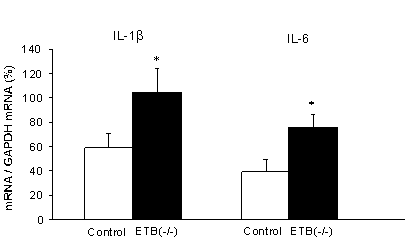 図7
図7
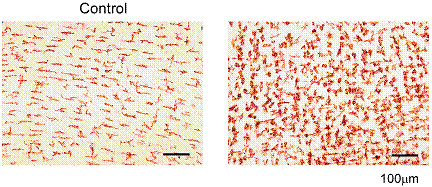 図8
図8
3)無神経節部ではなく膨大部の運動機能障害
さらに、ARの壁内神経叢を有する上部の消化管から輪状筋を摘出し、収縮反応を観察すると、リズムの乱れという特徴的な変化が認められた(図9)。また、自発性収縮の頻度も有意に低下していたが、炎症がみられない上部の十二指腸ではその様な変化は観察されなかった。さらに、ARのICCをc-kit抗体で染めてみると、対照に比べその数が有意に減少しており、形態変化も認められた(図10)。すなわち、腸管運動の減少は ICCの減少とそのネットワーク構造の変化によるものであると考えられた(20)。
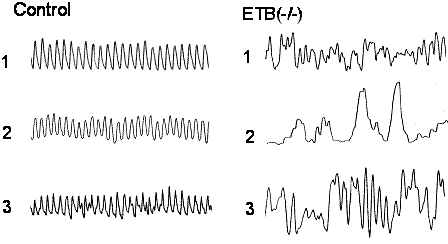 図9
図9
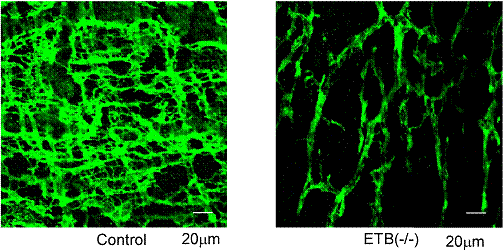 図10
図10
4)ヒルシュスプルング病腸管狭窄の3つの原因
これまで、ヒルシュスプルング病腸管狭窄の原因は、腸管蠕動の統合機能をつかさどる神経サーキットの欠除によると考えられてきた。しかし、本研究によりヒルシュスプルング病の進展により有神経節部にも炎症が起こり、ここに運動機能障害が生じて上部からの駆出力が減退することも一因となることが示された。重度のヒルシュスプルング病では無神経節腸管の切除術が施されるが、それでも以後の便秘傾向が完全に改善されるわけではなく、保存的な療法が取られる。この時、腸管炎症を如何にコントロールするか、すなわち腸内フローラのコントロールの重要性が示唆される。
ところで、ARの無神経節部分の腸管の平滑筋層は増殖して肥厚し、また平滑筋自身の収縮性も亢進し、このことも狭窄のもう1つの要因ともなっている(21)。この様に、ヒルシュスプルング病腸管狭窄の原因は、神経節が欠損していることに加え、少なくとも2つの二次的要因が関与しているものと考えられる(20, 21)。
4.まとめ
炎症を伴う腸疾患では粘膜の炎症が問題となるが、同時に消化管の運動機能が障害されることにも着目しなければならない。すなわち、粘膜の炎症が進行すると、この炎症反応は次第に筋層へも伝わる。筋層に炎症が及ぶと神経・ICC・平滑筋によって構築される運動系が障害される。この時、腸内フローラの変化が免疫反応を介して重要な役割を果たすと考えられる。すなわち、運動機能障害によって腸内フローラは乱れ、その結果管腔内に様々な有害物質が蓄積され、細菌の体内移行が進行し、粘膜の炎症が悪化する。粘膜炎症の悪化はさらに筋層炎症を悪化させ、悪循環に入るものと想像される。この時重要な役割を担うのが、筋層の自然免疫系をつかさどると考えられる常在型マクロファージである。
文 献
- Rennick, D.M., and Fort, M.M. 2000. Lessons from genetically engineered animal models. XII. IL-10-deficient (IL-10(-/-) mice and intestinal inflammation. Am J Physiol Gastrointest Liver Physiol 278:G829-833.
- Caenepeel, P., Janssens, J., Vantrappen, G., Eyssen, H., and Coremans, G. 1989. Interdigestive myoelectric complex in germ-free rats. Dig Dis Sci 34:1180-1184.
- Husebye, E., Hellstrom, P.M., and Midtvedt, T. 1992. Introduction of conventional microbial flora to germfree rats increases the frequency of migrating myoelectric complex. J. Gastrointest. Mot. 4:39-45.
- Husebye, E., Hellstrom, P.M., Sundler, F., Chen, J., and Midtvedt, T. 2001. Influence of microbial species on small intestinal myoelectric activity and transit in germ-free rats. Am J Physiol Gastrointest Liver Physiol 280:G368-380.
- Xia, Y., Hu, H.Z., Liu, S., Pothoulakis, C., and Wood, J.D. 2000. Clostridium difficile toxin A excites enteric neurones and suppresses sympathetic neurotransmission in the guinea pig. Gut 46:481-486.
- Nieuwenhuijs, V.B., van Duijvenbode-Beumer, H., Verheem, A., Visser, M.R., Verhoef, J., Gooszen, H.G., and Akkermans, L.M. 1999. The effects of ABT-229 and octreotide on interdigestive small bowel motility, bacterial overgrowth and bacterial translocation in rats. Eur J Clin Invest 29:33-40.
- Sanders, K.M., Ordog, T., and Ward, S.M. 2002. Physiology and pathophysiology of the interstitial cells of Cajal: from bench to bedside. IV. Genetic and animal models of GI motility disorders caused by loss of interstitial cells of Cajal. Am J Physiol Gastrointest Liver Physiol 282:G747-756.
- Sanders, K.M., Ordog, T., Koh, S.D., Torihashi, S., and Ward, S.M. 1999. Development and plasticity of interstitial cells of Cajal. Neurogastroenterol Motil 11:311-338.
- Sanders, K.M. 1996. A case for interstitial cells of Cajal as pacemakers and mediators of neurotransmission in the gastrointestinal tract. Gastroenterology 111:492-515.
- Ozaki, H., Kawai, T., Shuttleworth, C.W., Won, K.J., Suzuki, T., Sato, K., Horiguchi, H., Hori, M., Karaki, H., Torihashi, S., et al. 2004. Isolation and characterization of resident macrophages from the smooth muscle layers of murine small intestine. Neurogastroenterol Motil 16:39-51.
- Mikkelsen, H.B., Thuneberg, L., Rumessen, J.J., and Thorball, N. 1985. Macrophage-like cells in the muscularis externa of mouse small intestine. Anat Rec 213:77-86.
- Eskandari, M.K., Kalff, J.C., Billiar, T.R., Lee, K.K., and Bauer, A.J. 1999. LPS-induced muscularis macrophage nitric oxide suppresses rat jejunal circular muscle activity. Am J Physiol 277:G478-486.
- Torihashi, S., Ozaki, H., Hori, M., Kita, M., Ohota, S., and Karaki, H. 2000. Resident macrophages activated by lipopolysaccharide suppress muscle tension and initiate inflammatory response in the gastrointestinal muscle layer. Histochem Cell Biol 113:73-80.
- Hori, M., Kita, M., Torihashi, S., Miyamoto, S., Won, K.J., Sato, K., Ozaki, H., and Karaki, H. 2001. Upregulation of iNOS by COX-2 in muscularis resident macrophage of rat intestine stimulated with LPS. Am J Physiol Gastrointest Liver Physiol 280:G930-938.
- Turler, A., Schwarz, N.T., Turler, E., Kalff, J.C., and Bauer, A.J. 2002. MCP-1 causes leukocyte recruitment and subsequently endotoxemic ileus in rat. Am J Physiol Gastrointest Liver Physiol 282:G145-155.
- Ikadai H, F.H., Agematsu Y, et al. 1979. Observation of congenital aganglionosis rat(Hirschsprung's disease rat) and its genetical analysis. Cong Anom:31-36.
- Ikadai H, S.K., Fujita H, et al. 1981. Animal model of human disease: Hirschsprung's disease. Comput Pathol Bull. 13:3-4.
- Ceccherini, I., Zhang, A.L., Matera, I., Yang, G., Devoto, M., Romeo, G., and Cass, D.T. 1995. Interstitial deletion of the endothelin-B receptor gene in the spotting lethal (sl) rat. Hum Mol Genet 4:2089-2096.
- Hosoda, K., Hammer, R.E., Richardson, J.A., Baynash, A.G., Cheung, J.C., Giaid, A., and Yanagisawa, M. 1994. Targeted and natural (piebald-lethal) mutations of endothelin-B receptor gene produce megacolon associated with spotted coat color in mice. Cell 79:1267-1276.
- Suzuki, T., Won, K.J., Horiguchi, K., Kinoshita, K., Hori, M., Torihashi, S., Momotani, E., Itoh, K., Hirayama, K., Ward, S.M., et al. 2004. Muscularis inflammation and the loss of interstitial cells of Cajal in the endothelin ETB receptor null rat. Am J Physiol Gastrointest Liver Physiol.287, G638-G646.
- Won, K.J., Torihashi, S., Mitsui-Saito, M., Hori, M., Sato, K., Suzuki, T., Ozaki, H., and Karaki, H. 2002. Increased smooth muscle contractility of intestine in the genetic null of the endothelin ETB receptor: a rat model for long segment Hirschsprung's disease. Gut 50:355-360.